
形態特徵
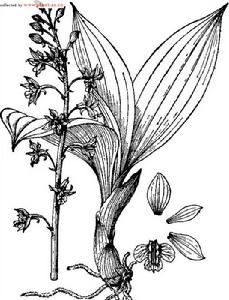 鉤距蝦脊蘭
鉤距蝦脊蘭根狀莖不明顯。假鱗莖短,近卵球形,粗約2厘米,具3--4枚鞘和3 4枚葉。假莖長5—18厘米,粗約1.5厘米。葉在花期尚未完全展開,橢圓形或橢圓狀披針形,長達33厘米,寬5.5—10厘米,先端急尖或銳尖,基部收狹為長達10厘米的柄,兩面無毛。花葶出自假莖上端的葉叢間,長達70厘米,高出葉層之外,密被短毛;花序柄常具1枚鱗片狀的鞘;鞘寬卵形,長約6毫米,先端漸尖,無毛;總狀花序長達32厘米,疏生多數花,無毛;花梗白色,連同綠色的於房長15--20毫米,弧形彎曲,密被短毛;花張開;萼片和花瓣在背面褐色,內面淡黃色;中萼片近橢圓形,長10--15毫米,寬5--6毫米,先端銳尖,基部收狹;具(3--)4--5條脈,無毛或背面疏被短毛;側萼片近似於中萼片,但稍狹;花瓣倒卵狀披針形,長9—13毫米,寬3--4毫米,先端銳尖,基部具短爪,具3--4條脈,無毛;唇瓣淺白色,3裂;側裂片稍斜的卵狀楔形,長約4毫米,基部約.1/9與蕊柱翅的外側邊’緣合生,先端圓鈍或斜截形;中裂片近方形或倒卵形,長約4毫米,寬3毫米,先端擴大,近截形並微凹,在凹處具短尖;唇盤上具4個褐色斑點和3條平行的龍骨狀脊;龍骨狀脊肉質,終止於中裂片的中部,其末端呈三角形隆起;距圓筒形,長10--13毫米,常鉤曲,末端變狹;外面疏被短毛;內面密被短毛;蕊柱長約4毫米,無毛;,蕊柱翅下延到唇瓣基部並與唇盤兩側的龍骨狀脊相連線;蕊喙2裂,裂片三角形,長約1毫米,先端尖牙齒狀,藥帽在前端驟然收狹而呈喙狀;花粉團棒狀,等大,長約2毫米,具明顯的花粉團柄;粘盤近長圓形,長約1毫米。花期3—5月。
產地分布
產安徽(黃山)、浙江(龍泉、遂昌、開化、鎮海等)、江西(永修、德興等)、台灣(北部山地)、湖北(鶴峰、合豐、興山一帶)、湖南(桑植、新寧、龍山)、廣東北部和西南部(乳源、信宜)、香港、廣西(鐘山、融水、全州)、四川西南部(雷波)、貴州(梵淨山、雷山、凱里)和雲南東南部(富寧)。生於海拔600—1500米的山谷溪邊、林下等陰濕處。模式標本采自台灣。
形態特徵:
假鱗莖圓錐形,粗約15毫米,具3--4枚鞘和3枚尚未展開的葉。假莖粗壯,長達 25厘米,粗1--2厘米。葉在花期尚未展開,橢圓形或橢圓狀披針形,長20--40厘米, 寬5—12厘米,先端急尖或銳尖,邊緣波狀,基部收狹為柄,背面被短毛;葉柄細長, 通常長20厘米以上,寬約5毫米。花葶出自假莖上端的葉間,長達80厘米,密被短 毛,近中部具1枚鱗片狀鞘;鞘膜質,寬卵狀披針形,長約1厘米;總狀花序長達30 厘米,疏生少數至20餘朵花;花苞片宿存,膜質,狹披針形,長7—10毫米,先端漸 尖,無毛;花梗綠色,連子房長1.5—2厘米,被短毛;花大,稍垂頭,具難聞氣味; 萼片和花瓣黃褐色;中萼片長圓狀披針形,長2.5—3.5厘米,中部寬6—10毫米,先 端急尖或漸尖,具5—7條脈,背面被短毛;側萼片相似於中萼片,等長,但稍狹,具 5條脈,背面被短毛;花瓣近橢圓形,長約2.3厘米,中部寬6毫米,先端漸尖,具3 脈,其兩側的主脈分枝,無毛;唇瓣檸檬黃色,基部具短爪,與整個蕊柱翅合生,3 裂;側裂片鐮狀長圓形,長約8毫米,兩側裂片先端之間相距約13毫米,先端斜截形; 中裂片狹倒卵狀長圓形,與側裂片約等寬,先端急尖或圓鈍並具短尖;唇盤上具3條平 行的波狀褶片;褶片先端高高隆起,中央1條較長,幾乎到達中裂片先端;距淺黃色, 纖細,長2--3毫米,外面被短毛,距口具白色絨毛;蕊柱長約5毫米,上端擴大,疏 被短毛;蕊喙2裂;裂片狹三角形,長1.7毫米,先端長漸尖;藥帽在前端收狹而呈喙 狀;花粉團稍扁的卵球形,長約2毫米;粘盤近圓形。 花期4—5月。
 |
| 鉤距蝦脊蘭 |
產地分布:
產廣西北部(龍勝)、四川(峨眉山)和雲南(富寧、廣南、蒙自、雙柏、景東、 景洪、維西、中甸一帶)。生於海拔1000--2600米的山地常綠闊葉林下和山谷溪邊。模 式標本采自雲南(蒙自)。
參考文獻:
Calanthe hancockii Rolfe in Kew Bull.197,1896,et J.Linn.Soc.Bot.36:25. 1903;Sehltr.In Fedde Repert.Sp.Nov.Beih.4:237.1919;S.Y.Hu in Quart. J.Taiwan Mus.25(3,4):202.1972;中國高等植物圖鑑5:723.圖8276.1976; 中國蘭花全書135.圖6—7(13,14).1998.
本組概述:
Sect.Calanthe----Calanthe sect.Eucalanthe J.J,Smith,Fl.Buit.6:202.1905.---Calanthe sect.Calothyrsus Schltr.In Fedde Repert.Sp.Nov.Beih.1:376.1912. 根狀莖不明顯;假鱗莖圓錐形或卵球形,較少為稍伸長的圓柱形;葉柄在與葉鞘相連線處無關節;花苞片宿存;花通常開放;蕊喙通常2--3裂。 本組模式種:Calanthe triplicata(Willem.) Ames (Orchis triplicata Willem.). 我國有42種,5個變種。
亞屬概述:
Subgen.Calanthe---Calanthe subgen Eucalanthe Schltr.In Fedde Repert.Sp.Nov.Beih.1:376.1912. 蕊柱無蕊柱足;唇瓣基部與蕊柱翅合生而形成長短不等的管。 本亞屬模式種:Calanthe triplieata (Willem.)Ames 我國有2個組48種5個變種。
本屬概述:
R.Br.in Bot.Reg.7:sub t.573.1821,nom.conserv. 地生草本。根圓柱形,細長而密被淡灰色長絨毛。根狀莖有或無。假鱗莖通常粗 短,圓錐狀,很少不明顯或伸長為圓柱形的。葉少數,常較大,少有狹窄而呈劍形或帶 狀,幼時席捲,全緣或波狀,基部收窄為長柄或近無柄,柄下為鞘,在葉柄與鞘相連線 處有一個關節或無,無毛或有毛,花期通常尚未全部展開或少有全部展開的。花葶出自 當年生由低出葉和葉鞘所形成的假莖上端的葉叢中,或側生於莖的基部,少有從去年生 無葉的莖端發出,直立,不分枝,下部具鞘或鱗片狀苞片,通常密被毛,少數無毛;總 狀花序具少數至多數花;花苞片小或大,宿存或早落;花通常張開,小至中等大;萼片 近相似,離生;花瓣比萼片小;唇瓣常比萼片大而短,基部與部分或全部蕊柱翅合生而 形成長度不等的管,少有貼生在蕊柱足末端而與蕊柱分離的,分裂或不裂,有距或無 距;唇盤具附屬物(胼胝體、褶片或脊突)或無附屬物;蕊柱通常粗短,無足或少數具短足,兩側具翅,翅向唇瓣基部延伸或不延伸;蕊喙分裂或不分裂;柱頭側生;花粉團 蠟質,8個,每4個為一群,近相等或不相等;花粉團柄明顯或不明顯,共同附著於1 個粘質物上。
全屬約150種,分布於亞洲熱帶和亞熱帶地區、紐幾內亞島、澳大利亞、熱帶非洲 以及中美洲。我國有49種及5變種,主要產長江流域及其以南各省區。
 |
| 鉤距蝦脊蘭 |
亞族概述:
Benth.In J.Linn.Soc.Bot.18:287.1881,"Bletieae". 地生、附生或腐生草本,具假鱗莖或球莖狀假鱗莖,較少僅具根狀莖;假鱗莖具 節,有1至數枚葉。葉螺鏇狀排列或二列,席捲式,摺扇狀,基部常有關節。花葶(或 花序)側生或有時頂生;花數朵至多朵,通常扭轉;萼片與花瓣離生;唇瓣有距或無 距;蕊柱不具或較少具蕊柱足;花葯俯傾;花粉團8個,4個成一群,蠟質,具或不具 花粉團柄,有時有粘盤或粘質物;柱頭凹陷。 本亞族模式屬:Bletia Ruiz et Pavon 共含17屬,分布於全球熱帶地區,向北到達北美洲和亞洲北部。我國有10屬。
本族概述:
Humb,Bonpl.EtKunth,Nov.Gen.Sp.Pl.1:269.1815.附生或較少地生或腐生草本,偶見攀援藤本;附生種類常有假鱗莖或肉質莖。葉基生或莖生,常為革質、紙質或肉質,罕有草質,少數無葉(腐生種類)。花變化較大;萼片合生或離生;唇瓣有距或無距;花葯以狹窄基部或花絲連線於蕊柱,常俯傾,兩藥室一般密接並藏於藥帽之內,花後常枯萎或脫落;花粉團一般4—8個,較少2個,蠟質或有時粒粉質,具或不具花粉團柄或粘盤,偶見粘盤柄;蕊喙一般位於柱頭上方。本族模式屬:EpidendrumL.共約340屬,全球均有分布,主要見於熱帶。我國有29個亞族含72屬。
亞科概述:
地生、附生或腐生草本,具根狀莖、塊莖或假鱗莖。葉莖生或基生,摺扇狀、對摺或具網狀脈,草質至革質。花序(或花葶)頂生或側生;花通常兩側對稱,有特化的唇瓣,少有例外,一般扭轉,具1室子房;蕊柱通常具1枚能育雄蕊;花粉粘合成團塊,稱花粉團;花粉團2--8個,粒粉質或蠟質,其一端常伸出花粉團柄並連線於粘盤,也有直接連線於粘盤或通過粘盤柄連線於粘盤的,較少既無花粉團柄又無粘盤柄與粘盤的;柱頭1或2,凹陷或凸出;蕊喙通常存在,大或小。果實通常為蒴果;較少呈莢果狀。種子通常兩端有延長的翅,較少無翅並有厚的外種皮。本亞科模式屬:OrchisL.共約690餘屬,全球均有分布。我國有4個族,42個亞族共167屬。
本科概述:
地生、附生或較少為腐生草本,極罕為攀援藤本;地生與腐生種類常有塊莖或肥厚的根狀莖,附生種類常有由莖的一部分膨大而成的肉質假鱗莖。葉基生或莖生,後者通常互生或生於假鱗莖頂端或近頂端處,扁平或有時圓柱形或兩側壓扁,基部具或不具關節。花葶或花序頂生或側生;花常排列成總狀花序或圓錐花序,少有為縮短的頭狀花序或減退為單花,兩性,通常兩側對稱;花被片6,2輪;萼片離生或不同程度的合生;中央1枚花瓣的形態常有較大的特化,明顯不同於2枚側生花瓣,稱唇瓣,唇瓣由於花(花梗和子房)作180°扭轉或90°彎曲,常處於下方(遠軸的一方);子房下位,1室,側膜胎座,較少3室而具中軸胎座;除子房外整個雌雄蕊器官完全融合成柱狀體,稱蕊柱;蕊柱頂端一般具藥床和1個花葯,腹面有1個柱頭穴,柱頭與花葯之間有1個舌狀器官,稱蕊喙(源自柱頭上裂片),極罕具2—3枚花葯(雄蕊)、2個隆起的柱頭或不具蕊喙的;
蕊柱基部有時向前下方延伸成足狀,稱蕊柱足,此時2枚側萼片基部常著生於蕊柱足上,形成囊狀結構,稱萼囊;花粉通常粘合成團塊,稱花粉團,花粉團的一端常變成柄狀物,稱花粉團柄;花粉團柄連線於由蕊喙的一部分變成固態粘塊即粘盤上,有時粘盤還有柄狀附屬物,稱粘盤柄;花粉團、花粉團柄、粘盤柄和粘盤連線在一起,稱花粉塊,但有的花粉塊不具花粉團柄或粘盤柄,有的不具粘盤而只有粘質團。果實通常為蒴果,較少呈莢果狀,具極多種子。種子細小,無胚乳,種皮常在兩端延長成翅狀。科的模式屬:OrchisL.全科約有700屬20000種,產全球熱帶地區和亞熱帶地區,少數種類也見於溫帶地區。我國有171屬1247種以及許多亞種、變種和變型。蘭科是一個進化而複雜的科,至今對科內各類群間親緣關係的了解仍然是十分初步的。在目前已發表的許多蘭科系統中,以R.Dressier(1981,1993)系統較為全面,但該系統對鳥巢蘭族(ridb.Neottieae)的處理是有欠缺的。幸而C.Seidenfaden等人在“TheOrehidsofIndochina”(OperaBot.114:5--502.1992)和“TheOrehidsofPeninsularMalaysiaandSingapore”(1992)中有較合理的處理。本書基本上採用了後者的框架,但將等級作了調整,而對亞族的組成則更多地參考Dressler系統(PhylogenyandClassifica-tionoftheOn:hidFamily,1993)。本書將蘭科分為3個亞科,蘭亞科之下分4個族。其中蘭族(Orchideae)與萬代蘭族(Vandeae)是比較自然的類群,而鳥巢蘭族(Neottieae)與樹蘭族(Epidendreae)則尚存在不少問題。在許多近代蘭科系統中,上述的4個族均被作為亞科,在亞科之下又有大量的族,在族之下則是亞族。應當說,大多數亞族的建立是比較客觀的。但族的組合則含有相當多的主觀性,甚至是勉強的拼湊。
因此,本書避繁就簡,在蘭亞科之下只承認4個族,族之下則包含較多的亞族。這樣不僅簡化了層次,而且更為客觀和實用。當然,這也只是權宜之計,有些問題仍然未能解決,如鳥巢蘭族與樹蘭族的特徵交叉過多,有些亞族較為龐雜等。這裡就不作詳細討論了。這裡要說明的是,本書所提供的“亞科與族的檢索表”和“蘭亞科分亞族檢索表”均系根據國產種類編寫而成的,未必完全符合族與亞族的全面特徵,在使用時務請加以注意。
