
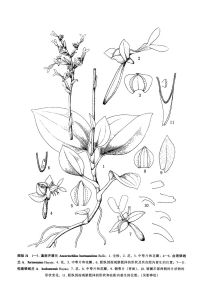
 恆春銀線蘭
恆春銀線蘭形態特徵
植株高約20厘米。根狀莖匍匐,伸長。莖肉質,圓柱形。葉2--4枚,卵形或卵圓形,長34厘米,寬23厘米,先端急尖,基部圓形,驟狹成柄,葉柄的基部具鞘;葉片上面呈絨毛狀,墨綠色,具白色美麗的網脈,葉背面帶紅色。總狀花序具5--6朵花,花莖長約15厘米,紅褐色,被毛,下部具2--3枚疏生的鞘狀苞片;花苞片卵形或卵狀披針形,與子房等長或較子房短,背面被毛;子房圓柱形,不扭轉,被毛,連花梗長約1.5厘米;花張開,不倒置(唇瓣位於上方);萼片紅褐色,背面被毛,中萼片卵形,凹陷呈舟狀,長5—5.5毫米,寬3--4毫米,先端驟狹、急尖;側萼片斜長橢圓形,長7—8毫米,寬3.5—3.8毫米,先端急尖;花瓣白色,鐮刀形,長5.5---6毫米,寬1.5—1.6毫米,先端漸尖且向內彎,與中萼片粘合呈兜狀;唇瓣白色,呈Y字形,基部具圓錐狀距,前部擴大成深2裂,裂片長橢圓形,長7—8毫米,寬3.5—3.7毫米,先端圓鈍,中部收狹成長約5毫米的爪,其兩側各具1枚長約5毫米、線狀披針形或三角形、其先端不裂或具2--3齒的片狀物;距微帶棕色,長6.5--7毫米,向上彎曲,與子房近呈直角伸展,其末端2裂,裂隙深約2毫米,其內側在靠近蕊柱基部處具2枚胼胝體;蕊柱長3—3.5毫米,前面兩側具2枚片狀附屬物;花葯卵形,長3.5毫米,寬1.5毫米,先端驟狹,漸尖;花粉團2個,長圓形,具長的花粉團柄,末端共同具1枚卵形的粘盤;蕊喙直立,叉狀2裂;柱頭2個,離生,位於蕊喙基部兩側。花期7—10月。產地分布
產我國台灣省,從北部至南部。生於海拔1500--1700米的常綠闊葉林下。模式標本采自台灣(恆春附近)。參考文獻
AnoectochiluskoshunensisHayata,Icon.Pl.Formos.4:104,fig.54.1914;台灣蘭科植物2:6070(圖).1977;台灣蘭科植物彩色圖鑑1:418.1977et2:3911990;台灣植物志5:876,圖1545.1978.——Odomochiluskoshunensis(Hayata)S.S.Ying,Col.Illust.Oreh.Fl.Taiwan1:64.1996.本屬概述
AnoectchilusBl.Bl,Bijdr.:411.20Sep-7Dec1825(‘Anecochilus’),nom.conserv.;Benth.EtHook.F,Gen.Pl.3:598.1833;Seidenf.InDanskBot.Ark.32(2):40.1978.——OdontochilusBl,1.C.地生蘭。根狀莖伸長,莖狀,匍匐,肉質,具節;節上生根。莖直立,或向上伸展,圓柱形,具葉。葉互生,常稍肉質,部分種的葉片上面具雜色的脈網或脈紋,基部通常偏斜,具柄。花大、中等大或較小,倒置(唇瓣位於下方)或不倒置(唇瓣位於上方),排列成疏散或較密集的、頂生的總狀花序;萼片離生,背面通常被毛,中萼片凹陷,舟狀,與花瓣粘合呈兜狀;側萼片常較中萼片稍長,基部圍繞唇瓣之基部;
花瓣較萼片薄,膜質,與中萼片近等長,常斜歪;唇瓣基部與蕊柱貼生,基部凹陷呈圓球狀的囊,囊小,被側萼片的基部包著,或延伸成圓錐狀的距,伸出於側萼片基部之外,下垂或上舉;唇瓣前部多明顯擴大成2裂,其裂片的形狀種種且叉開,罕前部略擴大,中部收狹成或長或短的爪,其兩側多具流蘇狀細裂條或具鋸齒而少為全緣的,基部囊或距的末端2淺裂或不裂,其內面中央具1枚縱向隔膜狀的龍骨狀的褶片或罕無,而在它的兩側各具1枚肉質、形狀種種、具短柄或無柄的胼胝體;蕊柱短,前面兩側常各具1枚縱向、平行的翼狀的附屬物或罕無;花葯2室;
花粉團2個,每個多少縱裂為2,棒狀,為具小團塊的粒粉質,具長或短的花粉團柄,共同具1個粘盤;蕊喙通常直立,叉狀2裂;柱頭2個,離生,凸出,位於蕊喙基部前方或基部的兩側,極罕2個柱頭緊靠一起而合生成較大的1個,位於蕊喙前面之下的正中央。本屬模式種:AnoectochilussetaceusBl.約有40餘種,分布於亞洲熱帶地區至大洋洲。我國有20種,2變種,產於西南部至南部。開唇蘭屬(AnoectochilusBl.)與齒唇蘭屬(OdontochilusBl.)兩者的區別很小,主要據前者的唇瓣基部延伸成圓錐狀的距,距較長,伸出於兩側萼片基部之外,不被側萼片基部包著,柱頭2個,位於蕊喙基部之兩側;後者唇瓣基部凹陷呈圓球狀囊,其囊小,藏於兩側萼片基部之內、被包著;柱頭2個(罕為1個),2個柱頭位於蕊喙基部兩側至基部的前方,確有過渡,極罕2個柱頭緊靠一起合生成1個,位於蕊喙前面之下正中央。多數學者主張將此兩屬合併。我們根據國產20種2變種植物花部結構的觀察研究,贊成C.Seidenfaden(1978年)將齒唇蘭屬(OdontochilusBl.)合併人開唇蘭屬(AnoectochilusBl.)中的見解。本屬植物部分種全草民間作藥用。
亞族概述
Subtrib.Goodyednae Klotz. Klotz.In Hayne,Arzbetgew 14:tab.24. 1846,"Goodyereae". 地生或罕有附生或腐生草本,常具近莖狀的圓柱形根狀莖和少數稍肉質的根。莖一 般較細弱,具葉。葉散生或簇生,席捲式,草質,常有種種美麗斑紋。總狀花序頂生, 具數花至多花;花一般扭轉,具較淺色澤;唇瓣基部通常凹陷成囊狀或有距,內有腺體 或毛,不同程度地與蕊柱基部合生,有時前端擴大,在囊狀基部與擴大前端之間具爪, 爪上常有流蘇或齒缺;花葯生於蕊柱背側,直立;花粉團2個或每個縱裂為2,花粉團 粒粉質,由許多可分的小團塊組成,具或不具花粉團柄,花粉團或花粉團柄從花葯頂端 伸出粘附於粘盤上;蕊喙直立,與花葯近等長;柱頭1或2個。 本亞族模式屬:Goodyera R.Br. 花約35屬,全球均有分布,主要見於熱帶,特別是亞洲熱帶地區。我國有13屬。
本族概述
Lindl,Coil.Bot.App.1821.地生或腐生草本,地下通常有長或短的根狀莖,極罕具塊莖,不具假鱗莖。葉基生或莖生;葉片通常扁平,罕有近圓筒形,少數無葉(腐生種類)。花兩側對稱或罕有近輻射對稱;唇瓣基部一般不具長距;花葯以狹窄基部或花絲連線於蕊柱,常枯萎或脫落;花粉團通常2個,罕有4個,粒粉質,為均勻的粒粉質或由可分的小團塊組成,具或不具花粉團柄和粘盤,若有花粉團柄則從花葯頂端伸出;蕊喙長或短,前者常直立並分裂。本族模式屬:NeottiaL.共約80屬,全球熱帶至溫帶均有分布。我國有8個亞族共45屬。
亞科概述
地生、附生或腐生草本,具根狀莖、塊莖或假鱗莖。葉莖生或基生,摺扇狀、對摺或具網狀脈,草質至革質。花序(或花葶)頂生或側生;花通常兩側對稱,有特化的唇瓣,少有例外,一般扭轉,具1室子房;蕊柱通常具1枚能育雄蕊;花粉粘合成團塊,稱花粉團;花粉團2--8個,粒粉質或蠟質,其一端常伸出花粉團柄並連線於粘盤,也有直接連線於粘盤或通過粘盤柄連線於粘盤的,較少既無花粉團柄又無粘盤柄與粘盤的;柱頭1或2,凹陷或凸出;蕊喙通常存在,大或小。果實通常為蒴果;較少呈莢果狀。種子通常兩端有延長的翅,較少無翅並有厚的外種皮。本亞科模式屬:OrchisL.共約690餘屬,全球均有分布。我國有4個族,42個亞族共167屬。
本科概述
地生、附生或較少為腐生草本,極罕為攀援藤本;地生與腐生種類常有塊莖或肥厚的根狀莖,附生種類常有由莖的一部分膨大而成的肉質假鱗莖。葉基生或莖生,後者通常互生或生於假鱗莖頂端或近頂端處,扁平或有時圓柱形或兩側壓扁,基部具或不具關節。花葶或花序頂生或側生;花常排列成總狀花序或圓錐花序,少有為縮短的頭狀花序或減退為單花,兩性,通常兩側對稱;花被片6,2輪;萼片離生或不同程度的合生;中央1枚花瓣的形態常有較大的特化,明顯不同於2枚側生花瓣,稱唇瓣,唇瓣由於花(花梗和子房)作180°扭轉或90°彎曲,常處於下方(遠軸的一方);子房下位,1室,側膜胎座,較少3室而具中軸胎座;除子房外整個雌雄蕊器官完全融合成柱狀體,稱蕊柱;
蕊柱頂端一般具藥床和1個花葯,腹面有1個柱頭穴,柱頭與花葯之間有1個舌狀器官,稱蕊喙(源自柱頭上裂片),極罕具2—3枚花葯(雄蕊)、2個隆起的柱頭或不具蕊喙的;蕊柱基部有時向前下方延伸成足狀,稱蕊柱足,此時2枚側萼片基部常著生於蕊柱足上,形成囊狀結構,稱萼囊;花粉通常粘合成團塊,稱花粉團,花粉團的一端常變成柄狀物,稱花粉團柄;花粉團柄連線於由蕊喙的一部分變成固態粘塊即粘盤上,有時粘盤還有柄狀附屬物,稱粘盤柄;花粉團、花粉團柄、粘盤柄和粘盤連線在一起,稱花粉塊,但有的花粉塊不具花粉團柄或粘盤柄,有的不具粘盤而只有粘質團。果實通常為蒴果,較少呈莢果狀,具極多種子。種子細小,無胚乳,種皮常在兩端延長成翅狀。科的模式屬:
OrchisL.全科約有700屬20000種,產全球熱帶地區和亞熱帶地區,少數種類也見於溫帶地區。我國有171屬1247種以及許多亞種、變種和變型。蘭科是一個進化而複雜的科,至今對科內各類群間親緣關係的了解仍然是十分初步的。在目前已發表的許多蘭科系統中,以R.Dressier(1981,1993)系統較為全面,但該系統對鳥巢蘭族(ridb.Neottieae)的處理是有欠缺的。幸而C.Seidenfaden等人在“TheOrehidsofIndochina”(OperaBot.114:5--502.1992)和“TheOrehidsofPeninsularMalaysiaandSingapore”(1992)中有較合理的處理。本書基本上採用了後者的框架,但將等級作了調整,而對亞族的組成則更多地參考Dressler系統(PhylogenyandClassifica-tionoftheOn:hidFamily,1993)。本書將蘭科分為3個亞科,蘭亞科之下分4個族。其中蘭族(Orchideae)與萬代蘭族(Vandeae)是比較自然的類群,而鳥巢蘭族(Neottieae)與樹蘭族(Epidendreae)則尚存在不少問題。在許多近代蘭科系統中,上述的4個族均被作為亞科,在亞科之下又有大量的族,在族之下則是亞族。應當說,大多數亞族的建立是比較客觀的。但族的組合則含有相當多的主觀性,甚至是勉強的拼湊。因此,本書避繁就簡,在蘭亞科之下只承認4個族,族之下則包含較多的亞族。這樣不僅簡化了層次,而且更為客觀和實用。當然,這也只是權宜之計,有些問題仍然未能解決,如鳥巢蘭族與樹蘭族的特徵交叉過多,有些亞族較為龐雜等。這裡就不作詳細討論了。這裡要說明的是,本書所提供的“亞科與族的檢索表”和“蘭亞科分亞族檢索表”均系根據國產種類編寫而成的,未必完全符合族與亞族的全面特徵,在使用時務請加以注意。
