
簡述
又稱“多孔動物”是一門最低等的多細胞動物。單體或群體,大小不一,小的幾毫米,大者可達2米。單體外形變化大,身體無固定形狀,有球形、樹枝形、管形、瓶形或圓柱形,體的中央為一中空的中央腔,體壁上穿有許多小孔,故又稱多孔動物。體壁由內外兩層細胞構成:外層為扁形細胞,起保護作用。內層為鞭毛細胞,起攝食與消化的作用。骨骼為矽質或鈣質骨針、骨絲。骨針起支撐身體的作用,可以保存化石。骨針的形狀有單軸型、雙軸型、三軸型或多軸型等,或可根據自中心點分出的數目而分為單射、雙射、多射等,也有的形狀不規則。海綿動物門可分為:普通海綿綱、玻璃海綿綱、鈣質海綿綱及骨海綿綱。海綿動物多為海生,淡水生活的很少。最早出現於前寒武紀,一直延續到現代,對地質學的研究具有很特殊的意義。
動物界中最原始、最低等的多細胞動物。個體形似瓶、壺、臼等,有時聯成群體。分布於海水、湖沼、溪流中,成體營固著生活。體壁上的許多小孔(又稱多孔動物)。游離的一端有大孔開口為出水口。一般有中央管腔。體壁由內外兩層細胞構成,外層細胞扁平,內層細胞生有鞭毛,多數具原生質領,稱做領細胞。由於領細胞鞭毛的有規律的擺動,引起水流通過體內的溝道。隨水流進入管腔的食物顆粒落入領內,被領細胞消化,不消化的東西隨水流從頂端出水口流出體外。在內外兩層細胞間還有一層為中膠層,其中有變形細胞、生殖細胞、海綿絲細胞,並具有支持作用的針狀骨胳,稱骨針。
外形特性
 海綿動物
海綿動物 海綿動物起源於5.7-5億年前的寒武紀;其中390屬已被確認源自白堊紀(1.35-0.65億年前)。形態各異,體形多數不對稱,少數輻射對稱,單生或群生;這種多孔濾食性生物體大多“無柄”地直接附著在基質上;它們沒有器官,也沒有明晰的組織,但有細胞的分化,其細胞種類非常複雜;它們的骨骼要么沒有含鈣或含矽的骨針,要么沒有有機的海綿硬蛋白纖維;有水溝系統;通常為雌雄同體,能進行有性繁殖和無性繁殖。
骨骼
海綿動物除了個別的科沒有骨骼之外,其他所有的種類都是具有骨骼的,骨骼是海綿動物的一個典型特徵,是用以分類的重要依據之一。海綿動物的骨骼有 骨針 (spicule)及 海綿絲 (sponginfiber)兩種類型,它們或散布在中膠層內,或突出到體表,或構成網架狀。骨骼具有支持及保護身體的功能。骨針的成分或是由碳酸鈣組成鈣質骨針,或是矽質骨針,其中還都可能包括微量的銅、鎂、鋅等離子。
骨針按其大小又可分為 大骨針(megasclere),構成支持身體的骨架;及 小骨針(microsclere),它散布在中膠層內,以支持體壁中的管道部分。小骨針僅存在於矽質海綿中。從形態上骨針可以分為多種,其中常見的有:
(1)單軸骨針(monaxons),即沿一個軸生長形成的骨針,軸或直或彎,軸的兩端或相似或不相似,末端或尖或具有其他改變;
 海綿動物
海綿動物 (2)四軸骨針(tetraxons),也稱 四放骨針(quadriradiate),這種骨針在一個平面上有四個放射端,但常因丟失一些放射端而變成三放、二放或一放型,三放骨針是鈣質海綿綱動物中最普通的一種骨骼;
(3)三軸骨針(triaxons),它的三個軸相互以直角癒合,因而呈 六放型(hexactinal),這種也常減少末端而改變放數,其末端可以彎曲、分枝、或具鈞、具結等變化而形成了多種形態;
(4)多軸骨針(polyaxons),由中心向外伸出多射,形成星壯,這種類型多見於小骨針。
不同種的海綿,各種骨針或彼此分離,或按一定結構形成疏鬆的或堅實的網架以支持身體,因此可根據骨針的類型、數量及排列而作為海綿動物分類的依據。海綿絲是一種纖維狀骨骼,它是由 硬蛋白(scleroprotein)組成,它們或單獨的存在於海綿動物體壁內,或與矽質骨針同時存在。許多小的矽質骨針埋在海綿絲中,形成有效地支持物。許多大型群體海綿常同時存在著這兩種骨骼。
海綿動物的骨針及海綿絲都是由中膠層中的變形細胞特化形成的造骨細胞所形成。單軸的鈣質骨針是由一個造骨細胞分泌形成,骨針形成時,造骨細胞核先分裂,並在雙核細胞的中心出現一個有機質的細絲,然後圍繞這一細絲沉積碳酸鈣,隨著骨針的逐漸增長,雙核細胞也分成兩個細胞,並分別加長骨針的兩端,最後形成一個單軸骨針。
同樣,三軸骨針是由三個造骨細胞聚集在一起,每個細胞也隨著有機質細絲的形成而分裂一次,形成六個細胞,碳酸鈣圍繞有機質細絲沉積癒合的結果形成了一個三軸型骨針。海綿絲是由許多造骨細胞聯合形成,先是由少數細胞形成分離的小段,然後再癒合成長的海綿絲。在尋常海綿綱動物中,這些海綿絲再相互聯結形成網狀骨架。
體型
從微小至2米長;其中最大的物種分布於 南極洲和 加勒比海。
形態
 海綿動物
海綿動物 所有海綿動物的結構都十分相似;它們簡單的體壁包括 表皮 (上皮)、 連線(連合)組織和 多種類型的細胞,其中包括能通過原生質的流動來移動(變形運動)的細胞(變形細胞)。這些變形細胞在其內部組織中游移,拉伸骨針並產生海綿硬蛋白絲。海綿動物並非完全不能移動,它們身體的主體能通過肌細胞的移動進行有限的活動,但在通常情況下,它們卻往往固定在同一地點紋絲不動。
海綿動物的身體柔軟,但許多觸摸起來卻很結實,這是因為它們的內骨骼是由堅硬的含鈣或含矽、桿狀或星狀的骨針和/或網狀蛋白質纖維即海綿硬蛋白所組成的,譬如浴海綿就是如此。有些物種的骨針可能穿透其海綿表面,一旦人們觸摸它們就會引起皮膚感染。海綿動物是濾食動物,它們濾取水中細小的碎石和細菌為食,分解其中的氧氣和有機物並將廢棄物排走。水通過海綿動物體表的細孔進入水溝系,並移動到順著環細胞或襟細胞這類有鞭毛的細胞排列的小室中;環細胞吸收通過在變形細胞間傳遞的食物顆粒,最後常通過其體表上火山狀的排水孔將水排出體外;水主要在環細胞鞭毛的作用下,穿過海綿動物的全身。
物種分類
海綿動物
六射海綿綱(六放海綿)
 海綿動物
海綿動物 約有600個物種,海生,一般棲息於潮位線之下,但在深海中更常見。其骨骼為複雜的矽質骨針,基本形態呈六放形。屬和種包括: 泡沫 海綿、 維納斯 花籃(EuplectellaAspergillum)、Holascus、圍線海綿。
鈣質海綿鈣質海綿綱
約有400個物種。海生。鈣質骨針的骨架如針形或三至四放形。
屬包括:Acyssa、Clathrina、Leucilla、白枝海綿、樽海綿。
尋常海綿尋常海綿綱
約有4,000個物種。海生和淡水生。其骨架或沒有矽質的骨針,或沒有有機的海綿硬蛋白纖維,或二者均無。當骨針存在時,則不為六放形。屬和種包括: 真海綿、穿貝海綿、 加勒比 海綿(Cribochalinavasulum)、刻盤海綿 、橘海綿、浴海綿(Hipposongiacommunis)、加勒比火海綿(Neofibularianolitangere)、管指海綿、沐浴角骨海綿(Spongiaofficinalis)、針海綿。
珊瑚海綿硬質海綿綱
 海綿動物
海綿動物 約有15個物種。海生,在熱帶的淺穴或深穴或其下的珊瑚上。骨架以鈣質為基礎,含有矽質骨針和有機纖維;形成覆蓋在鈣質基座上的薄薄一層海綿。屬包括: Ceratoporella、Stromatospongia。海綿動物在生殖發育中存在胚層逆轉現象。
身體結構
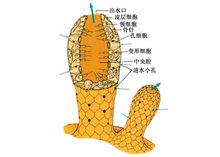 假胃腔
假胃腔 海綿動物身體的基本結構是由兩層細胞圍繞中央的一個空腔所組成。游離的一端有一個大的 出水口(osculum)使 中央腔(central cavity)與外界相通。構成海綿動物體壁的兩層細胞在不同的種類組成複雜程度不同的溝系,根據溝系可以將海綿動物的身體結構分為三種類型。
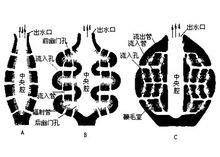
單溝型
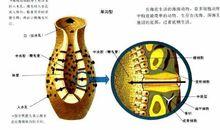
單溝型 (ascon type)是最原始,也是最簡單的體壁結構,種類很少,前述的白枝海綿就屬於這一類。單溝型海綿呈單體或群體,長度一般不超過10cm,群體中的個體輪廓明顯,每個個體均呈小管狀,出水口周圍有骨針包圍,中央腔寬闊,體壁由兩層細胞中間夾有 中膠質(mesoglea)所組成,外層細胞 稱皮層(dermal epithelium),主要是由一層 扁平細胞(pinacocytes)組成,它不同於其他動物的表皮層細胞,因為它們的來源和其他多細胞動物的表皮層不同,並且這種扁平細胞沒有基膜,細胞的邊緣可以收縮。
 水溝系分類
水溝系分類 許多扁平細胞同時收縮可以使身體變小。某些扁平細胞特化形成管狀,稱為 孔細胞 (porocyte),穿插在扁平細胞之間。孔細胞的外端與外界相通,內端與中央腔相通,孔細胞外端的小孔就是單溝型海綿動物體表的 進水小孔(ostia)或稱 流入孔(incurrent pore),所以它是細胞內孔,水由流入孔進入中央腔。孔細胞的收縮及舒張可以控制水的流入量。體壁的內層也稱 胃層(gastral epithelium),是由 領鞭毛細胞(也簡稱領細胞)組成,單溝型海綿的領細胞圍繞著整箇中央腔。領細胞呈卵圓形,其基部疏鬆的坐落在中膠層中,游離端伸出一根鞭毛,圍繞鞭毛的基部有一可伸縮的原生質領,是由許多分離的 微絨毛(microvilli)所組成。單溝型海綿通過領細胞鞭毛的擺動使水由 孔細胞(或稱入水小孔)流入,經中央腔再由出水口流出。
領細胞在形態上非常相似於原生動物門的領鞭毛蟲,因此有人認為海綿動物是由領鞭毛蟲進化而來。體壁的皮層與胃層之間是中膠層,它是一種含有蛋白質的膠狀透明基質,其中包括有游離的 變形細胞 (amoebocyte)及分散的 骨針(spicule)。變形細胞可以分化成不同的形態,例如有的變形細胞偽足細長分枝,彼此相連形成網狀,稱為 星芒細胞(collencyte),有人認為它是一種最原始的具有神經機能的細胞。另一種細胞較大,其細胞核也較大,有葉狀偽足,稱 原細胞 (archeocyte),這是一種未分化的細胞,除了本身具有吞噬及消化食物的機能外,它還可以轉化成具生殖功能的 生殖細胞(generative cell)、能分泌骨骼的 造骨細胞(scleroblast)、貯藏營養物質的 貯存細胞(thesocyte)、能分泌粘液的 腺細胞 (gland cell)等。
從上述可以看出單溝型海綿動物最大特徵是體壁結構簡單,其兩層細胞平直的包圍中央腔。由於中央腔寬闊,靠領細胞的鞭毛打動使流過身體的水流速度是緩慢的,代謝較低,所以單溝型海綿動物一般都是小型的。海綿動物在進化過程中通過體壁的褶疊增加了領細胞的數量及分布的表面積,同時減少了中央腔的體積,其結果是形成了雙溝型或復溝型的體壁,這樣就加速了水流過身體的速度,提高了代謝的能力,使動物也增大了體積。
雙溝型
雙溝型 (sycon type)是體壁褶疊的一種初步形式,例如 樽海綿(Scypha)、 毛壺(Grantia)等。雙溝型海綿皮層的扁平細胞褶向中膠層,形成多個平行排列的盲管,稱為 流入管(incurrent canal),流入管外端的開孔名為 流入孔(incurrentpore)。胃層的領細胞由中央腔向外端突出也形成多個穿插於流入管之間的盲管,稱為 鞭毛管(flagellated canal)或 鞭毛室,也稱為放射管 (radial canal),其內端的開孔稱為 後幽門孔(apopyle),結果流入管與鞭毛管相間排列形成了雙溝型的體壁。相鄰的流入管與鞭毛管之間也有小孔使兩管相通,這種小孔稱 前幽門孔(prosopyle)。
由於管道的出現,雙溝型的體壁加厚了,也由於領細胞褶入到鞭毛管中,中央腔壁上不再有領細胞,而是由扁平細胞包圍。雙溝型海綿的水流途徑是: 水→流入孔→流入管→前幽門孔→鞭毛管→後幽門孔→中央腔→出水口→體外。雙溝型海綿增加了領細胞層的面積,管道的增加及中央腔的縮小也加速了水流通過身體的速度。雙溝型海綿中,有些種類其皮層細胞及中膠層更發達,以致遮蓋了整個體表,形成了一層或薄厚不一的 外皮(cortex),結果出現了更多的流入孔,這樣可以增加體壁內的水壓,加速水在體內的流動。
復溝型
體壁進一步的褶疊複雜化就形成了 復溝型 (leucon type),大多數的海綿動物屬於這種類型,例如淡水海綿。復溝型結構的變化表現在:
(1)鞭毛管繼續向中膠層內褶入,以致形成了多個圓形的鞭毛室,例如 細芽海綿(Microciona)每平方毫米的體壁,鞭毛室可多達1000個;
(2)中膠層更發達,並與表皮細胞一起構成了眾多的 皮層孔(dermal pore)或 皮下腔(subdermal space);
(3)流入管分成許多小枝,然後再進入鞭毛室;
(4)中央腔進一步地縮小,最後被分枝的 出水管(excurrent canal)所代替。
復溝型海綿的水流途徑是: 水→皮層孔→皮下腔→流入管→前幽門孔→鞭毛室→後幽門孔→流出管→出水口→體外。在有些復溝型海綿,其前、後幽門孔延伸形成了 前、後幽門管(prosodus,aphodus),結構更複雜。因此復溝型海綿動物具有更大的領細胞表面積,體內有縱橫相通的管道,中央腔也進一步縮小變成了管狀,因此流經體內的水流量增多,水流速度加快。復溝型海綿的體積也都是較大型的,特別是在群體大型海綿中,我們僅能從許多出水口判斷出海綿個體的形態及大小,例如 磯海綿(Reniera)。淡水海綿的群體成團狀,已很難判斷出個體的形態了。
生活習性
 海綿動物
海綿動物 海綿動物雖然是多細胞動物中最簡單的一類,卻有一個龐大的家族,種數達10000多種,占所有海洋動物種數的1/15。由於海綿動物的體壁內長著具有支持作用的針狀骨骼,叫做骨針。通常根據骨針的性質,可以分為鈣質海綿和非鈣質海綿兩大類。體形最大的海綿動物是1909年曾在 巴哈馬群島撈獲的一隻,圍長為183厘米,剛出水重40千克,曬乾後的重量為5千多克。此外,在 安的列斯群島生活的一種海綿動物,身長106厘米,寬91.5厘米。海王星海綿也是體形較大的種類,剖面長120厘米,卻不太寬。最小的種類是白枝海綿,身高不過3毫米,體重僅有幾克,跟一粒芝麻一樣小。海綿動物的壽命也比較長,有的種類據說可以活幾百年。
海綿動物總是形單影隻地獨處一隅,凡是海綿動物棲居的地方就很少有其它動物前去居住。科學家分析這種現象形成的原因首先是海綿動物對那些貪食的動物沒有任何吸引力,它渾身的骨針和纖維使其它動物難以下咽,因此海綿動物的天敵不多。其次,海綿動物大多棲息在有海流流動的海底,而很多動物都難於在那樣的環境中生活。因為在那裡,它們的幼蟲或被水流沖走,或被海綿動物濾食。此外,海綿動物身上通常都有一股難聞的惡臭,這也是可能是其他動物不願與之為伍的原因之一。
捕食方法
 海綿
海綿 海綿動物是怎樣獲得食物的呢?它的捕食方法十分奇特,是用一種濾食方式。單體海綿很像一個花瓶,瓶壁上的每一個小孔都是一張“嘴巴”。海綿動物通過不斷振動體壁的鞭毛,使含有食餌的海水不斷從這些小孔滲入瓶腔,進入體內。
在“瓶”內壁有無數的領鞭毛細胞,由基部向頂端螺鏇式地波動,從而產生同一方向的引力,起到類似抽水機的泵吸作用。當海水從瓶壁滲入時,水中的營養物質,如動植物碎屑、藻類、細菌等,便被領鞭毛細胞捕捉後吞噬。經過消化吸收,那些不消化的東西隨海水從出水口流出體外。如果把石墨粉或幾滴墨水滴在飼養在水族箱中的活海綿動物的一側,過不了 多久瓶口(出水孔)處就會流出黑色的細流。隨著源源不斷的水流,細菌、硅藻、原生動物或有機碎屑也被攜入體內為領細胞俘獲供作營養。
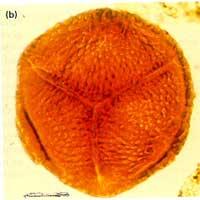 海綿動物
海綿動物 這種取食方式充分證明了它屬於濾食的異養動物。然而鞭毛的擺動是要耗能的。對營固著生活的海綿動物來說,從食物中獲得化學能來之不易。因此,海綿動物總是生活在有海流經過的海底,在千百萬年的進化過程中,完善了一套利用天然流體流動能的本領,從而節約了寶貴的食物化學能。一個高10厘米,直徑1厘米的海綿,一天內能抽海水22.5升,出水口處的水流速度可達5米/秒。這種高速離去的水流保證了從體內排出的廢物不再“回爐”。海綿動物正是有了濾食和節能的本領,才能在缺乏營養的熱帶珊瑚礁中和極地陸架區世代繁衍。
生長繁殖
生理特點
海綿動物是固著生活的,沒有移位的運動,僅能通過體表扁平細胞和孔細胞的收縮而略微改變身體的體積。許多雙溝型及復溝型海綿,在進水小孔及出水口的四周扁平細胞特化形成 類肌細胞(myocyte),類肌細胞的形狀及收縮性能有些類似平滑肌,它的收縮可以調節水流出入的速度,當在惡劣環境時,例如污水、暴露於空氣中等,類肌細胞可以關閉小孔或出水口,環境改善後,類肌細胞鬆弛,小孔又重新開放,這種反應是極為緩慢的,有時數分鐘後才能看到微小的變化;海綿動物的生理活動是依靠於通過身體的水流而進行,水流可以帶進食物及氧氣、帶走代謝廢物、生殖細胞,總之靠水流完成生長、發育及繁殖。
有人觀察過一種 白海綿(Leucandra),這是一種復溝系小型的鈣質海綿,流經出水口的水流速度是8.5cm/s,據估計直徑1cm、高10cm的白海綿大約有225萬個鞭毛室,每天有22.5L的海水流過身體。出水口狹窄,水流速度很快,而鞭毛室數量極多,其內水流緩慢,這樣便於領細胞有更多的時間從水中攝取食物及氣體交換。水在體內的流動是由領細胞的鞭毛打動所引起,鞭毛由基部向端部鏇轉運動,使水流進入鞭毛室。同一鞭毛室的鞭毛運動既不同步,也不互相協調,但鞭毛的方向都是指向後幽門孔。許多復溝型海綿在鞭毛室的出口處有一中央細胞,它的收縮可變動位置以調節水流的流量,甚至可以完全關閉後幽門孔而阻止水由鞭毛室流出。海綿動物取食各種有機物顆粒,食物的選擇主要是看有機物顆粒是否能進入入水孔。
有人用幾種海綿動物做實驗,結果發現食物中80%是細小的有機質顆粒,20%為細菌、鞭毛蟲類及其他極小的浮游生物。食物隨水流進入鞭毛室,水流速度在此減慢,有利於領細胞的捕食,領細胞領部的微絨毛可以粘著微小的食物顆粒,然後吞噬,較大的食物顆粒也可以被入水小管的扁平細胞所吞噬。食物進入領細胞後它可進行部分的細胞內消化,部分食物轉移到中膠層的變形細胞,在變形細胞中進行食物的消化。消化後的營養物或貯存在變形細胞內,或轉移到其他細胞,不能消化的食物殘渣仍由變形細胞運出。海綿動物沒有專營呼吸與排泄的細胞,而是當水流過身體時,大多數細胞均可與水接觸,各自獨立完成呼吸與排泄的機能。
許多淡水海綿,大多數細胞內具有一到幾個伸縮泡,這些伸縮泡像原生動物一樣,擔任著調節水與鹽份的平衡。海綿動物沒有神經結構,對刺激的反應常是局部的、緩慢的,對刺激反應的大小是依賴於刺激的強弱。信息物質的傳遞是通過中膠質中的擴散作用、游離變形細胞及固定細胞彼此的接觸而進行。尚未發現海綿動物具有電傳導。
生殖與發育
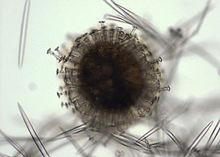 芽球
芽球 海綿動物的生殖有無性生殖和有性生殖。
無性生殖又分出芽和形成芽球兩種。 出芽(bud-ding)是由海綿體壁的一部分向外突出形成芽體,與母體脫離後長成新個體,或者不脫離母體形成群體。 芽球(gemmule)的形成是在中膠層中,由一些儲存了豐富營養的原細胞聚集成堆,外包以幾丁質膜和一層雙盤頭或短柱狀的小骨針,形成球形芽球。當成體死亡後,無數的芽球可以生存下來,渡過嚴冬或乾旱,當條件適合時,芽球內的細胞從芽球上的一個開口出來,發育成新個體。所有的淡水海綿和部分海產種類都能形成芽球。
有性生殖:海綿有些為 雌雄同體(monoecy),有些為 雌雄異體(dioecy)。精子和卵是由原細胞或領細胞發育來的。卵在中膠層里,精子不直接進入卵,而是由領細胞吞食精子後,失去鞭毛和領成為變形蟲狀,將精子帶入卵,進行受精。
 有性生殖
有性生殖 這是一種特殊的受精形式。就鈣質海綿來說受精卵進行卵裂,形成囊胚,動物極的小細胞向囊胚腔內生出鞭毛,另一端的大細胞中間形成一個開口,後來囊胚的小細胞由開口倒翻出來,裡面小細胞具鞭毛的一側翻到囊胚的表面。這樣,動物極的一端為具鞭毛的小細胞,植物極的一端為不具鞭毛的大細胞,此時稱為 兩囊幼蟲(amphiblastula),幼蟲從母體出水孔隨水流逸出,然後具鞭毛的小細胞內陷,形成內層,而另一端大細胞留在外邊形成外層細胞,這與其他多細胞動物原腸胚形成正相反(其他多細胞動物的植物極大細胞內陷成為內胚層,動物極小細胞形成外胚層),因此稱為 逆轉(inversion)。幼蟲遊動後不久即行固著,發育成成體。這種明顯的逆轉現象存在於鈣質海綿綱如 毛壺屬(Grantia)、 樽海綿屬(Sycon)、 白枝海綿屬(Leucosolenia)及尋常海綿綱的少數種類如 糊海綿屬(Oscurella)。其多數種類形成 實胚幼蟲(parenchymula larva),為另一種逆轉形式。
海綿的再生能力很強,如把海綿切成小塊,每塊都能獨立生活,而且能繼續長大。將海綿搗碎過篩,再混合在一起,同一種海綿能重新組成小海綿個體。有人將 細芽海綿屬(Microciona)與 穿貝海綿屬(Cliona)分別搗碎作成細胞懸液,兩者混合後,各按自己的種排列和聚合,逐漸形成了橘紅海綿與黃海綿。這對研究細胞如何結合很有意義。還有人用 細胞鬆弛素(cytochalasin)處理分離的海綿細胞,則能抑制其分離細胞的重聚合。
物種保護
在全球所有的海洋中,海綿動物的數量都十分巨大;在堅硬的基質上,它們更是多得驚人;相對而言,極少海綿動物能適應不穩定的沙地或泥沼的生存環境。它們的垂直生活領域從潮汐效應時水岸的最低處,並向下延伸至8,600米深的海洋深淵;矽質海綿中的淡水海綿科甚至能在全球的淡水湖泊和河流中生存。棲息於在潮標間的海綿動物通常只局限于海岸的一部分區域,即在空氣中暴露時間較短的那部分海岸。有些海綿也在高于海岸一點的地方出現,但僅限於棲息在被遮蔽的地方或背向太陽的岩石上。有些海綿動物一旦暴露在空氣中的時間略長就會死去,因此在大陸架的淺水域中,海綿的物種和個體數量都達到最大。
巨穴海綿常是更小動物的棲息處,這些小動物中的一部分對海綿動物無害,而另一部分則是寄生動物。許多海綿動物含有能進行光合作用的 單細胞藻類 (蟲綠藻)、 藍綠藻和可為海綿動物提供營養的共生細菌。 海蛞蝓 (海兔)、 石鱉 、 海星 (尤其是南極洲的)、 海龜和部分 熱帶魚都以海綿動物為食。在那些暴露在空氣中而不是躲藏在岩石下的熱帶海綿物種中,通常超過一半的都對魚類有毒。學者認為這是海綿動物對高密度魚類捕食的一種進化反應,大自然選擇了有害有毒的物質以防止魚類對海綿動物的捕食。部分有毒海綿動物體型巨大,如龐大的 加勒比海綿 (Cribochalinavasculum);而其他海綿動物觸摸起來十分危險,如 加勒比火海綿 (Neofibularianoltangere)――它們往往引起人類嚴重的灼燒感,並持續數小時。毒素能避免動物幼蟲和植物孢子附著在海綿動物表面,因此可能有益於保持其表面的清潔;有些海綿毒素還能防止其周圍無脊椎動物長的過大並導致海綿的窒息。
海綿毒素已被套用於神經刺激傳輸的研究中;它們顯示出作為生物性疏通減退劑的巨大潛力,並可作為鯊魚驅除物。
浴海綿在吸水和保持複雜的海綿硬蛋白纖維結構上有其自身的作用;這些纖維也具有彈性,能使水從海綿動物中被擠出來。許多海綿動物物種可被人們採集(特別是在 佛羅里達和 希臘的海岸),例如具有精細骨架網路的沐浴角骨海綿和骨架粗糙的馬海綿。它們生長於從低潮位到深海中的岩石底部,可通過船上的爪鉤或潛水者來獲取。海綿動物的加工方法簡單包括將其置於太陽下曬乾,使其軟組織腐爛,經過捶打和清洗只留下其海綿硬蛋白骨骼。通過切除栽培海綿動物的方法已經獲得成功,儘管這種方法在可行性上並不比人工合成更強。海綿動物含有各式各樣抗生物質、色素、諸如固醇之類的獨特化學物質、毒素、甚至抗發炎和治風濕的化合物。海若螺科中的穿貝海綿會使牡蠣殼變弱,從而造成經濟損失。這些海綿動物還能通過化學和機械兩種方式掘穴。
經濟價值
海綿動物對人有用的僅僅是海綿的骨骼,如浴海綿,因為海綿質纖維較軟,吸收液體的能力強,可供沐浴及醫學上吸收藥液、血液或膿液等用。其他有些種類纖維中或多或少的含有矽質骨骼,所以較硬,可用以擦機器等。天然產的海綿不夠用,有些地方還用人工方法繁殖,辦法是把海綿切成小塊,系在石架上,然後沉入海底,一般二三年即可成長。有些種類常長在牡蠣的殼上,會把殼封閉起來,造成牡蠣死亡。淡水海綿大量繁殖可以堵塞水道,這些對人都是有害的。有些淡水海綿要求一定的物理化學生活條件,因此可作為水環境的鑑別物。古生物學的研究表明,海綿的特殊沉積物對分析過去環境的變遷有意義。 對海綿的研究,年來發展也較快,不僅是研究海綿動物本身,而更重要的是用它作為研究生命科學基本問題的材料,如細胞和發育生物學等方面的一些基本問題,因此海綿動物對科學研究也有其特殊的意義。
海洋學相關知識(八)
| 海洋科學是研究海洋的自然現象、性質及其變化規律,以及與開發利用海洋有關的知識體系。下面讓我們來完善與海洋學相關的詞條,以此來更加深入的了解海洋學的相關知識。 |
