
形態特徵
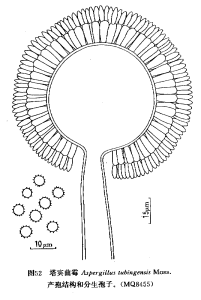 塔賓麴黴
塔賓麴黴 菌落在查氏瓊脂上生長迅速,25℃培養7天直徑50-75mm;平坦或近於平坦,邊緣具有白色的菌絲體;質地絲絨狀或稍帶絮狀;分生孢子結構大量,紅褐色至褐黑色;具或不具少量無色的滲出液;菌落反面帶黃色。分生孢子頭幼時球形至輻射形,直徑150-450μm,老時少有分裂成圓柱狀結構者;分生孢子梗發生於基質,孢梗莖(2000-)3000-5000(-5500)μm×12-20μm,老時黃色或黃褐色,壁平滑;頂囊球形或近球形,直徑(20-)40-70(-80)μm,全部表面可育;產孢結構雙層:梗基10-30(-35)μm×3-6μm,瓶梗8-10(-12)μm×2.5-3μm;分生孢子球形或近球形,直徑3-4μm,壁近於平滑至稍粗糙,老後出現縱向條紋。
菌落在查氏酵母膏瓊脂上生長很快,25℃7天直徑65-70mm,平坦,在邊緣有時具白色的菌絲體;質地絲絨狀;分生孢子結構大量,淡褐至褐黑色;無滲出液;菌落反面淡黃褐色至黃褐色。
菌落在麥芽汁瓊脂上生長最快,25℃7天直徑70-75mm;平坦;質地絲絨狀,具少量的不育性過度生長;分生孢子結構大量,淡褐色褐黑色;無滲出液;菌落反面黃褐色。
生境
土壤、哈密瓜、傘菌、牛糞。37℃生長。
中國分布
河北小五台山(C488);雲南景洪(C7666)、芒市(C6473);西藏定日(C2092)、吉隆(C1594);新疆(MQ8455)。
世界分布
印度。
模式產地
歐洲
特點
塔賓麴黴的特徵是具紅褐至褐黑色的菌落,較長的分生孢梗莖和較小的分生孢子。
Al-Musallam(1980)將本種內不同菌株分別處理為Aspergillusnigerv.Tiegh。和A.nigervar.intermediu(speg.)Al-Musallam的異名。Kozakiewicz(1989)則將本種作為A.niger的變種。kusters等(1991)研究黑麴黴的分類,根據核蛋白體DNA的限制性酶切片斷長度多態性分析結果認為本種是區別於A.niger的獨立的種。
有性型:石座菌屬PetromycesMalloch&Cain,can.J.Bot.50:2623.1972.
本組相當於Raper&Fennell(1965)分類系統的Aspergillusochraceus群。
分生孢子頭初為球形,老後呈緻密短柱形或裂成分叉的柱狀體,淺黃色、淡褐橄欖色、褐黃色或赭色;分生孢梗莖通常粗糙並帶褐色;頂囊大多為球形,全部表面可育;產孢結構大多為雙層;分生孢子較小,光滑或微粗糙;許多種能產生菌核。已知有兩個種產生堅硬如菌核樣的子座(stroma),內有閉囊殼,成熟極為緩慢。
模式種
AspergillusochraceusWilh.
Thom&Church(1926)建立了Aspergillusochraceus群,Raper&Fennell(1965)的專著中共承認了9個種,主要是依據菌核和孢子頭特徵區分的。以後不同的作者分別發表了9個新種。Subramanian(197l)根據Thom&Raper(1945)的討論認為AspergillusalutaceusBerk.&Curt.是A.ochraceus的最早的可用名稱而將後者降為異名。Gams等(1980)和Kozakiewicz(1989)都按此處理,並將A.ochraceus群改稱為A.alutaceus群;Kozakiewicz同時還把產有性型的A.alliaceus轉到A.wentii群。Chistensen(1982)研究本組的分類時,除以形態為基礎外,考慮到37℃是否生長這一特徵,並根據新增加的種對本組作了修訂,承認了15個種。我們認為Flavi組的A.leporisstates&Christ.與本組菌種更為相近而將之移入本組,同時將本組的A.sepultusTuth.&Christ.移入Wentii組(見Wentii組)。本組目前約有18個種,除少數幾個以外大多是罕見種,我國已知有8個。本組有些種可以轉化甾體化合物,有些種還可以產生抗生素與酶,可供工業利用,還有一些種產生赭麴黴毒素(ochratoxins),少數是昆蟲寄生菌。
盤點麴黴屬及其相關有性型的真菌
| 麴黴屬是叢梗孢目(Moniliales)叢梗孢科中的一屬。在自然界分布極廣,是引起多種物質霉腐的主要微生物之一。本期主要是盤點該屬及其相關。 |
