
形態特徵
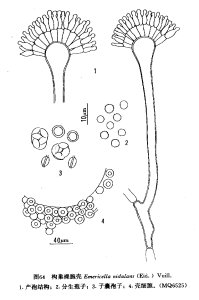 構巢裸胞殼
構巢裸胞殼菌落在查氏瓊脂上25℃7天直徑25mm左右,12-14天45-65mm,質地絲絨狀或絮狀,平坦或具不明顯的放射溝紋;分生孢子結構大量或較少,呈不同程度的綠色,近於不翡翠綠至水芹綠(JadeGreen-cressGreen,R.XXXI)或林肯綠至阿福花綠(LincolnGreen-asphodelGreen,R.XLI)乃至淺黃橄欖色(LightYellowishOlive,R.XXX);有部分閉囊殼散在其間,有的菌株產生大量的閉囊殼,由殼細胞所包圍,使表面顯現黃色團塊或絮狀顆粒結構,使菌落帶黃褐色,近於鹿褐色、淺黃褐色或古銅色(FawnColor,BuffyBrown,R.XL;IsabellaColor,R.XXX);滲出液多、少或無,呈淡紅褐至紫褐色;無氣味;菌落反面淡紫褐色至紫褐色,近於葡萄酒鹿褐色(VinaceousFawn,R.XL),稍擴散於基質。閉囊殼球形,暗紫紅色,直徑100-250(-300)μm,外面由殼細胞所包圍而呈現淡黃色;子囊近球形,8-12μm,一般在二周內成熟,有的則不成熟;子囊孢子雙凸鏡形,3.5-5μm×3.2-4μm,具兩個完整的赤道冠,具褶,寬約0.5-1μm,紫紅色,凸面光滑。分生孢子頭初為球形至放射形,直徑30-70μm,老後呈緻密短柱形,50-100μm×30-40μm;分生孢子梗生自基質或氣生菌絲孢梗莖大多較短(20-750-150(-300)μm×3.5-6.5μm,稍彎曲,帶茶褐色,壁光滑;頂囊為半球形,顏色與孢梗莖者相同,直徑(5.5-)8-16μm,上半部表面可育;產孢結構雙層:梗基4-8μm×2.5-3μm,有的異常菌株在頂囊上形成肥大的近於球形的梗基,瓶梗5-8μm×2-2.5μm;分生孢子球形,(2.5-)3-4(-4.5)μm,粗糙,偶而近於光滑;殼細胞大多為球形,直徑11-25μm,少數稍長或橢圓形18-40μm×12-24μm。
菌落在查氏酵母膏瓊脂上25℃7天直徑35-48mm,12-14天50-55mm;質地絲絨狀,中央部分氣生菌絲較多,平、薄或具不明顯的放射狀溝紋;分生孢子結構多,綠色,近於水芹綠色(CressGreen,R.XXXI),老後稍暗,近於暗草綠色(Kronberg'sGreen,R.XXXI);閉囊殼多或少;滲出液無色,呈小滴狀;菌落反面淡褐色到紫色,擴散於基質。
生境
菌落在麥芽汁瓊脂上生長更快,25℃7大直徑達40-70mm;質地絲絨狀,平、薄或具稀疏而不明顯的放射狀溝紋,中央有少量氣生菌絲,稍隆起;分生孢子結構大量,綠色,近於淺水芹綠至水芹綠色(LightCressGreen一CressGreen,R.XXXI)或阿福花綠至青豆綠(AsphodelGreen-poisGreen,R.XLI);閉囊殼多或少;有少量無色至褐色滲出液;菌落反面近於無色或黃褐色,老後變暗。
土壤,空氣,糧食,飼料,堆肥,皮革,木屑,牛糞,霉腐物,腦麴黴病患者等。37℃生長良好。
中國分布
北京(MQ20,MQ8644,MQ8685,MQ87l2,MQ8755,MQ8843);天津(MQ1941,MQ8661);河北小五台山(C44,C222,C322,C555,C590,Cl350);山西陽泉(MQ8438);遼寧鐵嶺(MQ5697);吉林長春(MQ1931);黑龍江北安(MQ8060)、大慶油田(MQ5705);江蘇揚州(MQ10606);浙江杭州(MQ5230,MQ10427)、金華(MQ5277);福建福州(MQ10123);山東濰坊(MQ5501)、泰山(MQ5480);河南鄭州(MQ8797)、潢川(MQ5636);不詳(MQ5016);湖北神農架(MQ6964,MQ7096,MQ7294,MQ7986,MQ9002,MQ9863);湖南長沙(MQ5389);廣東廣州(MQ1513)、東莞(MQ5599);雲南昆明(C4100,C4228,C85l7)、安寧(C4120)、思茅(C8101,C8377,C8424)、保山(C5372,C6314)、麗江(C4837,C5002,C5314)、景洪(C7800,C7987)、芒市(C6565,C6676),瑞麗(C6841,C6867)、大理(C5399,C5755,C5798);西藏定日(c2059)、吉隆(C1763)、亞東(MQ1602);陝西鹹陽(MQ4894)、富平(MQ4888);甘肅敦煌(MQ108l3);寧夏石嘴山(MQ6533)、陶樂(MQ6525)、賀蘭山(MQ6397,MQ6460,MQ6461);新疆呼圖壁(MQ30)、昭蘇(MQ4306)、托木爾峰(MQ4013);地點不詳(MQ8770)。
世界分布
廣布於全世界
模式產地
法國
本種特點
本種的明顯特徵是產生紫色的子囊孢子,具兩個有褶的雞冠狀突起,凸面光滑;產孢結構雙層;頂囊和孢梗基都帶有褐色。本種是研究遺傳學的理想材料,pontecorvo和Roper(1952)發現本種在無性繁殖過程中能形成雜合二倍體,從而發現了準性循環,已大量用於真菌遺傳學研究。它是研究真核基因表達和控制的最好的模型系統(Scazzochio,1992)。通過遺傳工程,可以利用本菌合成蛋白質(Martinellietal.,1992)。本菌還能引起人和動物致病;也能產生真菌毒素,主要是雜色麴黴素(sterigmatocystin)。
本菌最通用的名稱是構巢麴黴(Aspergillusnidulans),但作為無性型名稱是不合格的,因為原始描述包含了有性型,不符合國際植物命名法規第59條。(見命名的有關問題節),因此Samson和Gams(1985)提出了無性型的新名稱A.nidulellus。但由於AA.nidulans是不同領域工作者所熟知並長期廣泛使用的名稱,而且在遺傳學研究中積累了大量的文獻資料以及突變株和基因型,改用新名將引起混亂和不便,因此Frisvad等(1990),Kozakiewicz等(1992)提出了保留A.nidulans名稱的正式提案,業經命名委員會總會通過,作為保留名稱,繼續使用。
盤點麴黴屬及其相關有性型的真菌
| 麴黴屬是叢梗孢目(Moniliales)叢梗孢科中的一屬。在自然界分布極廣,是引起多種物質霉腐的主要微生物之一。本期主要是盤點該屬及其相關。 |
