
概述
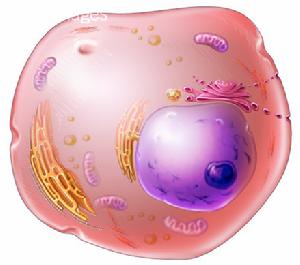 細胞質
細胞質細胞質(cytoplasm)是細胞質膜包圍的除核區外的一切半透明、膠狀、顆粒狀物質的總稱。含水量約80%。細胞質的主要成分為核糖體、貯藏物、多種酶類和中間代謝物、質粒、各種營養物和大分子的單體等,少數細菌還有類囊體、羧酶體、氣泡或伴孢晶體等。
細胞之中具有可辨認形態和能夠完成特定功能的結構叫做細胞器(organelles)。除細胞器外,細胞質的其餘部分稱為細胞質基質(cytoplasmicmatrix)或胞質溶膠(cytosol),其體積約占細胞質的一半。細胞質基質並不是均一的溶膠結構,其中還含有由微管、微絲和中間纖維組成的細胞骨架結構。
內膜系統介紹
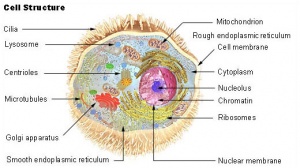 細胞質
細胞質內膜系統(endomembranesystem)是通過細胞膜的內陷而演變成的複雜系統。它構成各種細胞器(organelle),如內質網、線粒體、高爾基複合體、溶酶體等。這些細胞器均是互相分隔的封閉性區室,各具備一套獨特的酶系,執行著專一的生理功能。
內膜系統包括以下細胞器:
內質網
內質網(endoplasmicreticulum,ER)是扁平囊狀或管泡狀膜性結構,它們以分支互相吻合成為網路,其表面有附著核糖核蛋白s體者稱為粗面內質網(roughendoplasmicreticulum,RER),膜表面不附著核糖核蛋白體者稱為滑面內質網(smoothendoplasmicreticulum,SER),兩者有通連。
核糖核蛋白體附著在內質網上,其主要功能是合成分泌蛋白質(如免疫球蛋白、消化酶等),但也製造某些結構蛋白質(如膜鑲嵌蛋白質、溶酶體醇等)。粗面內質網分布於絕大部分細胞中,而在分泌蛋白旺盛的細胞(如漿細胞、腺細胞),粗面內質網特別發達,其扁囊密集呈板層狀,並占據細胞質很大一部分空間。一般說來,可根據粗面內質網的發達程度來判斷細胞的功能狀態和分化程度。
滑面內質網多是管泡狀,僅在某些組胞中很豐富,並因含有不同的酸類而功能各異,①類固醇激素的合成,在分泌類固醇激素的細胞中;滑面內質網膜上有合成膽固醇所需的酶系,在此合成的膽固醇再轉變為類固醇激素;②脂類代謝,小腸吸收細胞攝入脂肪酸、甘油及甘油一酯,在滑面內質網上酯化為甘油三酯,肝細胞攝取的脂肪酸也是在滑面內質網上被氧化還原酶分解,或者再度酯化;③解毒作用,肝細胞的滑面內質網含有參與解毒作用的各種酶系,某些外來藥物、有毒代謝產物及激素等在此經過氧化、還原,水解或結合等處理,成為無毒物質排出體外;④離於貯存與調節,橫紋肌細胞中的滑面內質網又稱肌漿網,其膜上有鈣泵,可將細胞質基質中的Ca2+泵入、貯存起來,導致肌細胞鬆弛,在特定因素作用下,貯存的Ca2+釋出,引起肌細胞收縮。胃底腺壁細胞的滑面內質網有氯泵,當分泌鹽酸時將CIˉ釋放,參與鹽酸的形成。
高爾基複合體
高爾基複合體(Golgicomplex)由扁平囊、小泡和大泡三部分組成,它在細胞中仿分布和數量依細胞的類型不同而異。扁平囊(saccule)有3-10層,平行緊密排列構成高爾基複合體的主體,它有一面常凸超稱生成面(formingface),另一面凹陷,稱成熟面(maturingface)扁平羹上有孔穿通,並朝向生成面。生成面附近有一些小泡(vesicle),直徑為40~80nm,是由附近粗面內質網芽生而來,將租面內質網中合成的蛋白質輕運到扁平囊,故小泡又稱運輸小泡。大泡(vacuole)位於成熟面,是高爾基複合體的生成產物,包括溶酶體、分泌泡等。溶酶體逐漸離開高爾基複合體而分散到細胞各部。分泌泡互相融合,其內容物電子密度增高,成為分泌顆粒。在蛋白質分泌旺盛的細胞中高爾基複合體發達。高爾基複合體對來自粗面內質網的蛋白質進行加工、修飾、糖化與濃縮,使之變為成熟的蛋白質,如在胰島B細胞中將前胰島素加工成為胰島素。高爾基複合體具有多種糖基轉移酶,許多蛋白質在此被糖化形成糖蛋白。此外,名種溶酶也在高爾基複合體濃聚形成初級溶酶體。
溶酶體
溶酶體(lysosome)為有膜包裹的小體,內含多種酸性水解酶,如酸性磷酸酶、組織蛋白酶、膠原蛋白酶、核糖核酸酶、葡萄糖苷酸和脂酶等,能分解各種內源性或外源性物質。它們的最適ph為5.0。不向細胞中的溶酶體不盡相同,(但均含酸性磷酸酶,故該酶為溶酶體的標誌酶。按溶酶體是否含有被消化物質(底物)可將其分為初級溶酶體(primarylysosme)和次級溶酶體(secondarylysosome)。
1、初級溶酶體:也稱原溶酶體(protolysosome)。一般呈圓形或橢圓形,直徑多介於25~50nm近年發現亦有長桿狀或緩狀溶酶體。其內容物呈均質狀,電子密度中等或較高不含底物。在少數細胞,如破骨細胞和炎症部位的中性粒細胞,溶酶體酶可被釋放到細胞外發揮水解作用。
2、次級溶酶體:也稱吞噬性溶酶體(phagolysosome)是由刺級溶酶體和將被水解的各種吞噬底物融合而構成,因此其體積較大,形態多樣,內容物為非均質狀。根據其作用廢物的來源不同,分為自噬性溶酶體和異噬性溶酶體。自噬性溶酶體(autophagolysosome)的作用底物是內源性的,即來自細胞內的衰老和崩解的細胞器或局部細胞質等。異噬性溶酶體(heterophagolysosome)的作用底物是經由細胞的吞飲或吞噬而被攝入細胞內的外源性物質,是溶酶體與吞噬體融合而成,多見於吞噬了細菌的中性粒細胞和吞噬了異物的巨噬細胞。並噬性溶酶體與自噬性溶酶體中的底物有的被分解為單糖、胺基酸等小分子物質,它們可通過溶酶體膜進入細胞質基質,被細胞利用;有的則不能被消化(如塵埃、金屬顆粒等異物、衰老細胞器的某些類脂成分),它們殘留於溶酶體中,當溶酶體酶活性耗竭,溶酶體內完全由殘留物占據,則稱之為殘餘體(residualbody)。在哺乳動物,殘餘體滯留在細胞中,常見的殘餘體有脂褐素顆粒和髓樣結構。均由自噬性溶酶體演化而來。脂褐素顆粒(lipofuscingranule)為不規則形,由電子密度不同的物質及脂滴構成,在光鏡下呈褐色,多見於神經細胞、心肌細胞、肝細胞及分泌類固醇激素的細胞,並隨年齡增長而增多。髓樣結構(myelinfigure)的內部為大量板層排列的膜,可能因膜性成分消化不全所致。初級溶酶體與吞飲小泡或其它小泡融合形成多泡體(multivesicularbody),其外有界膜,內含很多低電子密度小泡,基質具有酸性鱗酸酶活性。
線粒體
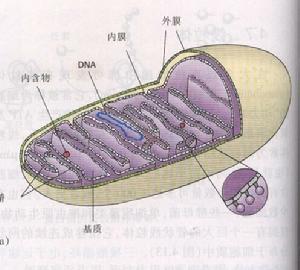 細胞質
細胞質線粒體(mitochondria)常為桿或橢圓形,橫徑為0.5~1ηm長2~6ηm但在不同類型激胞中線粒體的形狀、大小和數量差異甚大。電鏡下,線粒體具有雙層膜,外膜光滑,厚6~7nm,膜中有2~3nm小孔,分子量為1萬以內的物質可自由通過;內膜厚5~6nm,通透性較小。外膜與內膜之間有約8nm。膜間腔,或稱外腔。由膜向內摺疊形成線粒體嵴(mitochohdrialcrista),嵴之間為嵴間腔,或稱內腔,充滿線粒體基質。基質中常可見散在的,直徑25~50nm。電子緻密的嗜餓酸基質顆粒(matrixgranule),主要由磷脂蛋白組成,並含有鈣、鎂、磷等元素。基質中除基質顆粒外還含有脂類、蛋白質、環狀DNA分子核糖體。線粒體嵴膜上有許多有柄小球體,即基粒(elementaryparticle),其直徑為8~10nm,它由頭、柄和基片三部分組成。球形的頭與柄相連而突出於內膜表面,基片鑲嵌於膜脂中。
基粒中含有ATP合成酶,能利用呼吸鏈產生的能量合成ATP,並把能量貯存於ATP中。細胞生命活動所需能量的約95%由線粒體以ATP的方式提供,因此,線粒體是細胞能量代謝中心,線粒體嵴實為擴大了內膜面積,故代謝率高,耗能多的細胞。嵴多而密集大部分細胞的線粒體嵴為板層狀。桿狀線粒體的嵴多與其長軸垂直排列,圓形線粒體的嵴多以周圍向中央放射狀排列;在少數細胞,主要基分泌類固醇激素的細胞(如腎上腺皮質細胞等),線粒體峭多呈管狀或泡狀;有些細胞(如肝細胞)的線粒體兼有板層狀和管狀兩種。
線粒體另一個功能特點是可以合成一些蛋白質。目前推測,線上粒體中合成的蛋白質約占線粒體全部蛋白的10%,這些蛋白疏水性強,和內膜結合在一起。線起體合成蛋白質均是按照細胞核基因組的編碼輯導合成。如果沒有細胞核遺傳系統,線粒體RNA則不能表達。因此表明線粒體會成蛋白質的半自主性。
關於線粒體形成的機制,較普遍接受的看法是,線粒體依靠分裂而進行增殖。線粒體的發生過程可分為兩個階段,在第一階段中,線粒體的膜進行生長和複製,然後分裂增殖。第二階段包括線粒體本身的分化過程,建成能夠行使氧化磷酸化功能的機構。線粒體生長和分化階段分別接受兩個獨立遺傳系統的控制,因此,它不是一個完全自我複製的實體。
過氧化物酶體
過氧化物酶體(peroxisome)又稱微體(microbody),是有膜包裹的圓形小體,直徑為0.2~0.4μm,多見於肝細胞與腎小管上疫細胞。在人其內容物為低電子密度的均質狀;在某些動物尚含電子緻密的核心,是尿酸氫化酶的結晶。過氧化物體含有40多種酶,不同細胞所含酶的種類不同,但過氧化氫酶則存在所有細胞的過氧化物酶體中。各種氧酶能使相應的底物氧化,在氧化底物過程中,氧化酶使氧還原成過氧化氫,而過氧化氫酶能使過氧化氫還原成水。這種氧化反應在肝、腎細胞中是非常重要的。
核糖體
核糖體(ribosme)是由核糖體RNA(rRNA)和蛋白質組成的橢圓形緻密顆粒,並非膜性結構,(因屬細胞器,故在此敘述)顆粒大小約為15nm×25nm。核糖體由一個大亞基與一個小亞基構成。大亞基合兩條rRNA與約40個相關蛋白質分子,並有一條中央曾;小亞基含一條rRNA與約40個相關蛋白質分子,非功能狀態的核糖體單個存在。當一定數量(3~30)的核糖體由一條mRNA細絲穿行於它們的大、小亞基之間把它們串聯起來,則成為功能狀態的多核糖體(polyribosome),電鏡下呈串珠狀或花簇狀。核糖體能將mRNA所含的核苷酸密碼翻譯為胺基酸序列,即肽鏈合成的肽鏈從大亞基中央管釋出,肽鏈可進一步聚合形成白質細胞質基質中的游離核糖體(freeribosome)合成細胞自身的結構蛋白,如細胞骨架蛋白細胞基質中的酶類等,供細胞代謝、增殖和生長需要。因此,在旺盛增殖中的細胞游離核糖體極多。於內質網膜表面的附著核糖體(attachedribosome)除合成結構蛋白外,主要合成分泌性蛋白。核糖體豐富的細胞,光鏡下胞質呈嗜鹼性。
細胞骨架
 細胞質
細胞質細胞的特定形狀以及運動等,均有賴於細胞質內蛋白質絲織成的網狀結構——細胞骨架(cytoskeleton)。細胞骨架是由微管、微絲、中間絲和微梁網組成。
微管
微管(microtubule)是細而長的中空圓柱狀結構。管徑約15nm,長短不等,常數根平行排列。微管由微管蛋白(thbulin)聚合而成。微管蛋白單體為直徑約5nm的球形蛋白質,它們串連成原纖維,13條原纖維縱向平行排列圍成微管。微管有單微管、二聯微管和三聯做管三種類型。細胞中絕大部分微管為單微管,在低溫、Ca2+和秋水仙素作均下易解聚為微管蛋白,故屬於不穩定微管。二聯微管主要位於纖毛與精子鞭毛中,三聯微管參與構成中心體和基體,均為穩定微管。
微管具有多種功能。微管的支架作用可保持細胞形狀,如血小板周邊部的環行微管使其呈雙凸圓盤狀,神經細胞的微管支撐其突起,如果加入秋水仙素使微管解聚,則血小板變圓,神經細胞突起縮回。微管參與細胞的運動,如細胞分裂時,由微管組成的紡錘體可使染色體向兩極移動,如果加入秋水仙素則分裂停止於中期,纖毛和鞭毛的擺動、胞吞和胞吐作用、細胞內物質的運送都需要微管參與。
微絲
微絲(microfilament)廣泛存在於多種細胞中,微絲常成群或成束存在,在一些高度特化的細胞(如肌細胞),它們能形成穩定的結構,但更常見的是形成不穩定的束或複雜的網。它們可根據細胞周期和運動狀態的需要,改變其在細胞內的形態和空間位置,並能夠根據在細胞的不同狀態而聚合或解聚。
分布於肌細胞和非肌細胞中的微絲分細絲和粗絲兩種。細絲(thinfilament)直徑約6nm,長約lμm,主要由肌動蛋白(actin)組成,故又稱肌動蛋白絲(actinfilament),通常所說的微絲指此而言。細胞鬆弛素B能使細絲解聚,從而抑制細胞運動;粗絲(thickfilament)直徑側10~15nm,長約1.5μm,主要由肌球蛋白(myosin)組成,故又稱肌球蛋白絲(myosinfilament)。
微絲是肌細胞內的恆定結構。在橫紋肌細胞內;細絲與粗絲以一定比例(約為2:1)有規則排列成肌原纖維,其收縮機制已明確。平滑肌細胞內細絲與粗絲之比約為15:1,二者的排列不規則。非肌細胞中一般只能看到細絲,粗絲可能因存在時間短暫,或於電鏡標本製備過程中解聚為肌球蛋白,難於觀察到。在某些因素作用下,非肌細胞中的微絲迅速解策為其結構蛋白;在相反因素作用下,結構蛋白又裝配成微絲。其中細絲交聯成網以構成細胞骨架的一部分,並維持細胞質基質的膠質狀態;細絲與粗絲的局部相互作用能引發運動。在活躍運動的細胞(主要在細胞質周邊部)或細胞局部(如偽足),以及需察機械支持的部位(如微絨毛),都有豐富的微絲。因此,微絲除具有支持作用外,還參與細胞的收縮、變形運動、細胞質流動、細胞質分裂以及胞吞、胞吐過程。
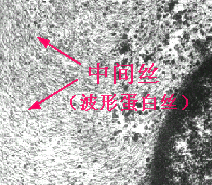
中間絲
中間絲(intermediatefilament)又稱中等纖維,直徑約為8~11nm,介於細絲與粗絲之間,因而得名。中間絲可分為五種,各由不同蛋白質構成。在成體中絕大部分細胞僅含有一種中間絲,故具有組織特異性,且較穩定。五種中間絲的形態相仿,難於分辨。但用免疫組織化學方法則能將它們區分,從而可進一步分析細胞的類型。
1)、角質蛋白絲(keratinfilament):分布於上皮細胞,在復層扁平上皮細胞內尤其豐富,常聚集成束,又稱張力絲(tonofilament)。張力絲附著於橋粒(一種細胞連線),能加固細胞間的連線。張力絲除起支持作用外,還有助於保持細胞的韌性和彈性。
2)、結蛋白絲(desminfilament):分布於肌細胞,在橫紋肌細胞內,結蛋白絲所形成的細網連線相鄰肌原纖維並使肌節位置對齊;在Z膜股處,細網包圍肌原纖維並與細胞膜連線。在平滑肌細胞內,結蛋白絲連線在密體與密斑之間形成立體網架,並與肌動蛋白絲相連。總之,結蛋白絲作為肌細胞的細胞骨架網,發揮固定和機械性整合作用。
3)、波形蛋白絲(vimentinfilament):主要存在於成纖維細胞和來自胚胎間充質的細胞。在少數含有兩種中間絲的細胞中,波形蛋白絲是其中的一種,波形蛋白絲主要在核周形成網架,對核起機械性支持,並穩定其在細胞內的位置。
4)、神經絲(neurofilament):存在於神經細胞的胞體與突起中,由神經絲蛋白組成,與微管共同構成細胞骨架,並協助物質運輸。
5)、神經膠質絲(neurogialfilament):主要存在於星形膠質細胞內,由膠質原纖維酸性蛋白組成,多聚集成束,交織走行於胞體,並伸入突起內。
微梁網
微梁網(microtrabecularlattict)是用超高壓電鏡等技術在完整細胞中觀察到的由直徑3~6nm的纖維交織形成的立體網架。有人認為它是一種鑲嵌在其它纖維系統中的微梁格線。也有人認為,它是微管、微絲和中間絲系統緊密聯繫和交錯相插,或是某些被磨損的細胞骨架所顯示的圖像。總之,它仍是一個有爭議的結構。
中心體
中心體(centrosome)多位於細胞核周圍,由一對互相垂直的中心粒(centriole)構成。中心粒呈是短圓筒狀,長0.5μm直徑為外0.2μm,由9組三聯微管與少量電子緻密的均質狀物構成其壁。相鄰的三聯微管相互斜向排列,狀如風車鏇翼。在壁外側有時可見9個球形的中心粒衛星(centriolarsatellite)。大小約70nm。在細胞分裂時,以中心粒衛星為起點形成紡錘體,參與染色體的分離(詳見"細胞周期")。有纖毛或鞭毛的細胞,中心粒形成基體,參與微管組的形成。
包涵物
是細胞質中本身沒有代謝活性,卻有特定形態的結構。有的是貯存的能源物質,如糖源顆粒、脂滴;有的是細胞產物,如分泌顆粒、黑素顆粒;殘餘體也可視為包涵物。
糖原顆粒
(glycogen granule)是細胞貯存葡萄糖的存在形式,於PAS反應時呈紅色。電鏡下,其電子密度高,無膜包裹,並呈兩種類型:β顆粒,直徑為20~30nm,形狀不規則,分散存在。多見於肌細胞;α顆粒,是β顆粒的聚合體,呈花簇狀,大小不一,多見於肝細胞。
脂滴
 細胞結構
細胞結構(fat drop)是細胞貯存脂類的存在形式,內含甘油三酯、脂肪酸、膽固醇等。脂滴在脂肪細胞中最多,
其次為分泌類固醇激素的細胞。在前者,常常一個脂滴即占據細胞的絕大部分空間;在後者,則多是小的球狀。在普通光鏡標本製備過程中,脂滴被二甲苯、乙醇溶解而遺留大小不等的空泡。電鏡下,脂滴無膜包裹,多是低或中等電子密度,與所含脂肪酸的不飽和程度有關。
分泌顆粒
(secretory granule)常見於各種腺細胞、內含酶、激素等生物活性物質。分泌顆粒的形態、大小及在細胞內的分布位置因細胞種類而異,但都有膜包裹。
遺傳特性
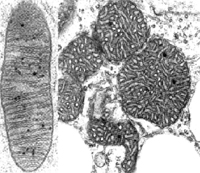 細胞質
細胞質細胞質遺傳的物質基礎是細胞質中的DNA,細胞質遺傳在實踐中的套用很廣泛。細胞質遺傳的概念:由細胞質基因所決定的遺傳現象和遺傳規律,也稱為非孟德爾遺傳,核外遺傳。
細胞質遺傳的特性
1、後代的表型象母親(又叫母系遺傳,偏母遺傳);
2、不遵循孟德爾遺傳,後代不出現一定的比例;
3、正交和反交後代的表型不同。
細胞質遺傳的機制
精卵結合中形成的合子父母雙親所提供的遺傳物質不均等,在雜種受精卵的原生質體中,核來自於父母雙方,而細胞質卻幾乎完全來自其母親(精子受精時胞質很少甚至不能進入卵細胞中)。在細胞分裂過程中,細胞質基因呈現不均等分配,因此細胞質遺傳不遵循孟德爾定律。
物質基礎
線粒體基因組(mtDNA)
葉綠體基因組(ctDNACpDNA)
細胞共生體基因組
細菌質粒基因組
非細胞器基因組
細胞器基因組
細胞質基因組
葉綠體基因組
1、細胞核遺傳與細胞質遺傳的區別
1)、細胞核和細胞質的遺傳物質都是DNA分子,但是分布的位置不同。細胞核遺傳的遺傳物質在細胞核中,細胞質遺傳的遺傳物質在細胞質中。
2)、細胞核和細胞質的遺傳橋樑都是配子,但是細胞核遺傳雌雄配子的核遺傳物質相等,而細胞質遺傳物質主要存在於卵細胞中。
3)、細胞核和細胞質的性狀表達主要通過體細胞進行的。核遺傳物質的載體(染色體)有均分機制,進行均分遵循遺傳規律;細胞質遺傳物質的載體(具有DNA的細胞器)沒有均分機制,而是隨機的。
4)、細胞核遺傳時,正反交相同.細胞質遺傳時,F1的性狀均與母本相同,即母系遺傳。
2、線粒體和葉綠體是半自主性細胞器
研究發現,線粒體和葉綠體中除有DNA外,還有RNA(mRNA、tRNA、rRNA),核糖體等。說明這兩種細胞器都具有獨立進行轉錄和翻譯的功能,也就是說,線粒體和葉綠體都具有自身轉錄RNA和翻譯蛋白質的體系。但迄今為止,人們發現葉綠體只能合成13種蛋白質,線粒體能夠合成的蛋白質也只有60多種,而參與組成線粒體和葉綠體的蛋白質卻分別上千種。這說明,線粒體和葉綠體中自身編碼,合成的蛋白質並不多,它們中的絕大多數蛋白質是由核基因編碼,在細胞質核糖體上合成的。也就是說,線粒體和葉綠體的自主程度是有限的,它們對核遺傳系統有很大的依賴性。因此,線粒體和葉綠體的生長和增殖是受核基因組及自身的基因組兩套遺傳信息系統控制的,所以它們都被稱為半自主性細胞器。
主要作用
細胞質是進行新陳代謝的主要場所,絕大多數的化學反應都在細胞質中進行。同時它對細胞核也有調控作用。
細胞質(cytoplasm)細胞質膜以內、細胞核以外的部分。由均質半透明的胞質溶膠(cytosol)和細胞器及內含物組成。胞質溶膠約占細胞體積1/2,含無機離子(如K+、Mg2+、Ca2+等)、脂類、糖類、胺基酸、蛋白質(包含酶類及構成細胞骨架的蛋白)等。骨架蛋白與細胞形態和運動密切相關,被認為對胞質溶膠中酶反應提供了有利的框架結構。絕大部分物質中間代謝(如醣酵解作用、胺基酸、脂肪酸和核苷酸代謝)和一些蛋白的修飾作用(如磷酸化)在胞質溶膠中進行。懸浮在胞質溶膠中的細胞器,有具界膜的和無界膜的,它們參與了細胞的多種代謝途徑。內含物則是在細胞生命代謝過程中形成的產物,如糖原、色素粒、脂肪滴等。
細胞質研究史
1665 英國人Robert Hook用自己設計與製造的顯微鏡(放大倍數為40-140倍,圖1-1)觀察了軟木(櫟樹皮)的薄片,第一次描述了植物細胞的構造,並首次用拉丁文cella(小室)這個詞來稱呼他所看到的類似蜂巢的極小的封閉狀小室(實際上只是觀察到到纖維質的細胞壁)。
1672,1682英國人Nehemaih Grew出版了兩卷植物顯微圖譜,注意到了植物細胞中細胞壁與細胞質的區別。
1846德國人H. von Mohl研究了植物原生質,發表了“identifies protoplasm as the substance of cells”。
1865 德國人J. von Suchs 發現葉綠體。
1868 英國人T. H. Huxley 在愛丁堡作題為“生命的物質基礎”(the physical basis of life)的演講報告時首次把原生質的概念介紹給了英國公眾。
1882 德國人E. Strasburger 提出細胞質(cytoplasm)和核質(nucleoplasm)的概念。
1898 義大利人C. Golgi 用銀染法觀察高爾基體。
50年代就有人看到葉綠體中有呈孚爾根反應的顆粒存在,推測其中可能有DNA。
1962年,Ris和Plant用電鏡觀察衣藻,玉米等植物的葉綠體超薄切片,發現在基質中電子密度較低的部分有20.5nm左右的細纖維.用DNA酶處理時消失,證明是DNA。
生物化學術語(一)
| 我們學習了很多的生物化學的專業術語。現在,我把它們都整理在一起,可以使我們更加全面、完整的學習,認識,了解它們。 |
