
組成與分類
組成
飽和脂肪酸(saturatedfattyacid):不含有—C=C—雙鍵的脂肪酸。
不飽和脂肪酸(unsaturatedfattyacid):至少含有—C=C—雙鍵的脂肪酸。
必需脂肪酸(occentialfattyacid):維持哺乳動物正常生長所必需的,而動物又不能合成的脂肪酸,如亞油酸,亞麻酸。
三脂醯苷油(triacylglycerol):又稱為甘油三酯。一種含有與甘油脂化的三個脂醯基的酯。脂肪和油是三脂醯甘油的混合物。
磷脂(phospholipid):含有磷酸成分的脂。如卵磷脂,腦磷脂。
鞘脂(sphingolipid):一類含有鞘氨醇骨架的兩性脂,一端連線著一個長連的脂肪酸,另一端為一個極性和醇。鞘脂包括鞘磷脂,腦磷脂以及神經節苷脂,一般存在於植物和動物細胞膜內,尤其是在中樞神經系統的組織內含量豐富。
鞘磷脂(sphingomyelin):一種由神經醯胺的C-1羥基上連線了磷酸毛里求膽鹼(或磷酸乙醯胺)構成的鞘脂。鞘磷脂存在於在多數哺乳動物動物細胞的質膜內,是髓鞘的主要成分。
卵磷脂(lecithin):即磷脂醯膽鹼(PC),是磷脂醯與膽鹼形成的複合物。
腦磷脂(cephalin):即磷脂醯乙醇胺(PE),是磷脂醯與乙醇胺形成的複合物。
脂質體(liposome):是由包圍水相空間的磷脂雙層形成的囊泡(小泡)。
分類
自然界約有40多種不同的脂肪酸,它們是脂類的關鍵成分。許多脂類的物理特性取決於脂肪酸的飽和程度和碳鏈的長度,其中能為人體吸收、利用的只有偶數碳原子的脂肪酸。脂肪酸可按其結構不同進行分類,也可從營養學角度,按其對人體營養價值進行分類。按碳鏈長度不同分類。它可被分成短鏈(含2~4個碳原子)脂肪酸;中鏈(含6~12個碳原子)脂肪酸和長鏈(含14個以上碳原子)脂肪酸肪酸三類。人體內主要含有長鏈脂肪酸組成的脂類。
飽和度 脂肪酸分離設備
脂肪酸分離設備它可分為飽和與不飽和脂肪酸兩大類。其中不飽和脂肪酸再按不飽和程度分為單不飽和脂肪酸與多不飽和脂肪酸。單不飽和脂肪酸,在分子結構中僅有一個雙鍵;多不飽和脂肪酸,在分子結構中含兩個或兩個以上雙鍵。隨著營養科學的發展,發現雙鍵所在的位置影響脂肪酸的營養價值,因此又常按其雙鍵位置進行分類。雙鍵的位置可從脂肪酸分子結構的兩端第一個碳原子開始編號。常從脂肪酸,並以其第一個雙鍵出現的位置的不同分別稱為ω-3族、ω-6族、ω-9族等不飽和脂肪酸,這一種分類方法在營養學上更有實用意義。
營養角度非必需脂肪酸是機體可以自行合成,不必依靠食物供應的脂肪酸,它包括飽和脂肪酸和一些單不飽和脂肪酸。而必需脂肪酸為人體健康和生命所必需,但機體自己不能合成,必須依賴食物供應,它們都是不飽和脂肪酸,均屬於ω-3族和ω-6族多不飽和脂肪酸。過去只重視ω-6族的亞油酸等,認為它們是必需脂肪酸,比較肯定的必需脂肪酸只有亞油酸。它們可由亞油酸轉變而成,在亞油酸供給充裕時這兩種脂肪酸即不至缺乏。自發現ω-3族脂肪酸以來,其生理功能及營養上的重要性越來越被人們重視。ω-3族脂肪酸包括麻酸及一些多不飽和脂肪酸,它們不少存在於深海魚的魚油中,其生理功能及營養作用有待開發與進一步研究。必需脂肪酸不僅為營養所必需,而且與兒童生長發育和成長健康有關,更有降血脂、防治冠心病等治療作用,且與智力發育、記憶等生理功能有一定關係。
人體組織脂肪酸含量| 人體各組織脂肪中脂肪酸的含量 | |||||||
| 脂肪酸 | 肝 | 心肌 | 脂肪組織 | 主動脈 | 心肌 | 心肌 | |
| 線粒體 | 微粒體 | ||||||
| 12:0 | 0.8 | 1 | |||||
| 14:0 | 1.1 | 4.6 | 4.6 | 3.1 | 4.1 | 4.9 | 4.7 |
| 14:1 | 0.6 | 0.7 | 0.4 | 1.1 | |||
| 15:0 | 0.2 | 0.3 | 0.5 | ||||
| 16:0 | 28.2 | 26.4 | 23.9 | 26.6 | 29.9 | 30.3 | 28.1 |
| 16:1 | 2.5 | 8.6 | 7.2 | 5.4 | 5.7 | 10.5 | 12.7 |
| 17:0 | 0.6 | 0.4 | 0.5 | ||||
| 17:1 | 0.3 | 0.5 | |||||
| 18:0 | 7.4 | 6.1 | 7.5 | 6.3 | 8.4 | 5.8 | 5.3 |
| 18:1 | 50.5 | 45.6 | 46.2 | 44.5 | 45.5 | 42.5 | 40.8 |
| 18:2 | 7.1 | 5.5 | 6.3 | 9.7 | 5.3 | 5.2 | 5 |
| 18:3(20:0) | 0.4 | 0.7 | 1 | 0.9 | |||
| 20:1 | 0.5 | 0.9 | 1.1 | 1 | |||
| 20:2 | 1.3 | ||||||
| 20:3 | 0.2 | 0.6 | |||||
| 20:4 | 1.3 | 1.2 | |||||
| 20:5 | 0.4 | ||||||
常用食用油脂中主要脂肪酸的組成
| 常用食用油脂中主要脂肪酸的組成(食物中脂肪總量的質量分數) | |||||
| 食用油脂 | 飽和脂肪酸 | 不飽和脂肪酸 | 其他脂肪酸 | ||
| 油酸(C18:1) | 亞油酸(C18:2) | 亞麻酸(C18:3) | |||
| 可可油 | 93 | 6 | 1 | ||
| 椰子油 | 92 | 0 | 6 | 2 | |
| 橄欖油 | 10 | 83 | 7 | ||
| 菜籽油 | 13 | 20 | 16 | 9 | 42* |
| 花生油 | 19 | 41 | 38 | 0.4 | 1 |
| 茶油 | 10 | 79 | 10 | 1 | 1 |
| 葵花籽油 | 14 | 19 | 63 | 5 | |
| 豆油 | 16 | 22 | 52 | 7 | 3 |
| 棉籽油 | 24 | 25 | 44 | 0.4 | 3 |
| 大麻油 | 15 | 39 | 45 | 0.5 | 1 |
| 芝麻油 | 15 | 38 | 46 | 0.3 | 1 |
| 玉米油 | 15 | 27 | 56 | 0.6 | 1 |
| 棕櫚油 | 42 | 44 | 12 | ||
| 米糠油 | 20 | 43 | 33 | 3 | |
| 文冠果油 | 8 | 31 | 48 | 14 | |
| 豬油 | 43 | 44 | 9 | 3 | |
| 牛油 | 62 | 29 | 2 | 1 | 7 |
| 羊油 | 57 | 33 | 3 | 2 | 3 |
| 黃油 | 56 | 32 | 4 | 1.3 | 4 |
註:*主要為芥酸
β-氧化
肝和肌肉是進行脂肪酸氧化最活躍的組織,其最主要的氧化形式是β-氧化。此過程可分為活化,轉移,β-氧化共三個階段。
脂肪酸的活化
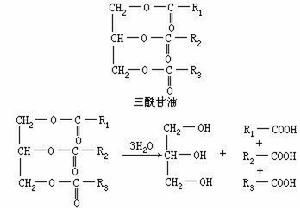 甘油三酯
甘油三酯和葡萄糖一樣,脂肪酸參加代謝前也先要活化。其活化形式是硫酯——脂肪醯CoA,催化脂肪酸活化的酶是脂醯CoA合成酶(acyl CoA synthetase)。
活化後生成的脂醯CoA極性增強,易溶於水;分子中有高能鍵、性質活潑;是酶的特異底物,與酶的親和力大,因此更容易參加反應。
脂醯CoA合成酶又稱硫激酶,分布在胞漿、線粒體膜和內質網膜上。胞漿中的硫激酶催化中短鏈脂肪酸活化;內質網膜上的酶活化長鏈脂肪酸,生成脂醯CoA,然後進入內質網用於甘油三酯合成;而線粒體膜上的酶活化的長鏈脂醯CoA,進入線粒體進入β-氧化。
脂醯CoA進入線粒體
催化脂肪酸β-氧化的酶系線上粒體基質中,但長鏈脂醯CoA不能自由通過線粒體內膜,要進入線粒體基質就需要載體轉運,這一載體就是肉毒鹼(carnitine),即3-羥-4-三甲氨基丁酸。
長鏈脂肪醯CoA和肉毒鹼反應,生成輔酶A和脂醯肉毒鹼,脂肪醯基與肉毒鹼的3-羥基通過酯鍵相連線。催化此反應的酶為肉毒鹼脂醯轉移酶(carnitine acyl transferase)。線粒體內膜的內外兩側均有此酶,系同工酶,分別稱為肉毒鹼脂醯轉移酶I和肉毒鹼脂醯轉移酶Ⅱ。酶Ⅰ使胞漿的脂醯CoA轉化為輔酶A和脂肪醯肉毒鹼,後者進入線粒體內膜。位於線粒體內膜內側的酶Ⅱ又使脂肪醯肉毒鹼轉化成肉毒鹼和脂醯CoA,肉毒鹼重新發揮其載體功能,脂醯CoA則進入線粒體基質,成為脂肪酸β-氧化酶系的底物。
長鏈脂醯CoA進入線粒體的速度受到肉毒鹼脂醯轉移酶Ⅰ和酶Ⅱ的調節,酶Ⅰ受丙二醯CoA抑制,酶Ⅱ受胰島素抑制。丙二醯CoA是合成脂肪酸的原料,胰島素通過誘導乙醯CoA羧化酶的合成使丙二醯CoA濃度增加,進而抑制酶Ⅰ。可以看出胰島素對肉毒鹼脂醯轉移酶Ⅰ和酶Ⅱ有間接或直接抑制作用。飢餓或禁食時胰島素分泌減少,肉毒鹼脂醯轉移酶Ⅰ和酶Ⅱ活性增高,轉移的長鏈脂肪酸進入線粒體氧化供能。
氧化反應過程
脂醯CoA線上粒體基質中進入β氧化要經過四步反應,即脫氫、加水、再脫氫和硫解,生成一分子乙醯CoA和一個少兩個碳的新的脂醯CoA。
第一步:脫氫(dehydrogenation)反應由脂醯CoA脫氫酶活化,輔基為FAD,脂醯CoA在α和β碳原子上各脫去一個氫原子生成具有反式雙鍵的α,β-烯脂肪醯輔酶A。
第二步:加水(hydration)反應由烯醯CoA水合酶催化,生成具有L-構型的β-羥脂醯CoA。
第三步:脫氫反應是在β-羥脂肪醯CoA脫飴酶(輔酶為NAD+)催化下,β-羥脂肪醯CoA脫氫生成β酮脂醯CoA。
第四步:硫解(thiolysis)反應由β-酮硫解酶催化,β-酮酯醯CoA在α和β碳原子之間斷鏈,加上一分子輔酶A生成乙醯CoA和一個少兩個碳原子的脂醯CoA。
上述四步反應與TCA循環中由琥珀酸經延胡索酸、蘋果酸生成草醯乙酸的過程相似,只是β-氧化的第四步反應是硫解,而草醯乙酸的下一步反應是與乙醯CoA縮合生成檸檬酸。長鏈脂醯CoA經上面一次循環,碳鏈減少兩個碳原子,生成一分子乙醯CoA,多次重複上面的循環,就會逐步生成乙醯CoA。
從上述可以看出脂肪酸的β-氧化過程具有以下特點:首先要將脂肪酸活化生成脂醯CoA,這是一個耗能過程。中、短鏈脂肪酸不需載體可直接進入線粒體,而長鏈脂醯CoA需要肉毒鹼轉運。β-氧化反應線上粒體內進行,因此沒有線粒體的紅細胞不能氧化脂肪酸供能。β-氧化過程中有FADH2和NADH+H+生成,這些氫要經呼吸鏈傳遞給氧生成水,需要氧參加,乙醯CoA的氧化也需要氧。因此,β-氧化是絕對需氧的過程。
生理意義
脂肪酸β-氧化是體內脂肪酸分解的主要途徑,脂肪酸氧化可以供應機體所需要的大量能量,以十六個碳原子的飽和脂肪酸軟脂酸為例,其β-氧化的總反應為:
CH3(CH2)14COSCoA+7NAD++7FAD+HSCoA+7H2O——→8CH3COSCoA+7FADH2+7NADH+7H
7分子FADH2提供7×2=14分子ATP,7分子NADH+H+提供7×3=21分子ATP,8分子乙醯CoA完全氧化提供8×12=96個分子ATP,因此一克分子軟脂酸完全氧化生成CO2和H2O,共提供131克分子ATP。軟脂酸的活化過程消耗2克分子ATP,所以一克分子軟脂酸完全氧化可淨生成129克分子ATP。脂肪酸氧化時釋放出來的能量約有40%為機體利用合成高能化合物,其餘60%以熱的形式釋出,熱效率為40%,說明機體能很有效地利用脂肪酸氧化所提供的能量。
脂肪酸β-氧化也是脂肪酸的改造過程,機體所需要的脂肪酸鏈的長短不同,通過β-氧化可將長鏈脂肪酸改造成長度適宜的脂肪酸,供機體代謝所需。脂肪酸β-氧化過程中生成的乙醯CoA是一種十分重要的中間化合物,乙醯CoA除能進入三羧酸循環氧化供能外,還是許多重要化合物合成的原料,如酮體、膽固醇和類固醇化合物。
產品用途
 丁苯橡膠
丁苯橡膠主要用於生產丁苯橡膠乳化劑、ABS樹脂、非離子表面活性劑、高級香皂、日用化妝品、洗滌劑、工業脂肪酸鹽、塗料、油漆等。
發展前景
從行業發展周期的角度,我國脂肪酸及其衍生物行業已經進入行業的成熟期。這一時期的脂肪酸及其衍生物市場增長率不高,技術上漸漸成熟,競爭狀況基本穩定,買方市場形成,行業盈利能力下降,新產品和產品的新用途開困難,行業進入壁壘很高。
根據我國脂肪酸及其衍生物工業的發展現狀及相關工業的發展要求,並借鑑世界脂肪酸及其衍生物工業的技術進展和發展趨勢,21世紀,我國脂肪酸及其衍生物工業發展的指導思想應當是:大力發展高性能、低污染的脂肪酸及其衍生物品種,並促進脂肪酸及其衍生物生產向專業化方向發展。
安全信息
危險品標誌:C,T
危險類別碼:34-37-23/24-22
安全說明:26-36/37/39-45
危險品運輸編號:UN33169/PG3
維生素:20世紀偉大的發現
| 維生素的發現是20世紀的偉大發現之一。1897年,C.艾克曼在爪哇發現只吃精磨的白米即可患腳氣病,未經碾磨的糙米能治療這種病。並發現可治腳氣病的物質能用水或酒精提取,當時稱這種物質為“水溶性B”。1906年證明食物中含有除蛋白質、脂類、碳水化合物、無機鹽和水以外的“輔助因素”,其量很小,但為動物生長所必需。 |
