
詳細描述
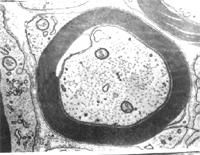 有髓神經纖維
有髓神經纖維有髓神經纖維(myelinated nerve fiber)由軸突(或樹突)、髓鞘、神經膜構成。髓鞘(myelin sheath)及神經膜(neurolemma)呈鞘狀包裹在軸突的周圍。在軸突的起始部無髓鞘包裹,稱此部為起始段(initial segment)起始段遠側的軸突部分,髓鞘呈節段包捲軸突,形似藕節,其間斷部位,軸膜裸露,,可發生膜電位變化,稱此部位為神經纖維節(node of nerve fiber),又稱郎飛結(Ranvier node)。兩個相鄰結之間的一段,稱結間體(internode),長約0.5~lmm,它是由一個施萬細胞所形成的髓鞘及其周圍的神經膜構成。施萬細胞核呈長橢圓形,位於髓鞘邊緣的少量胞質內。髓鞘主要是由類脂質和蛋白質所組成,稱為髓磷脂(myelin)在常規染色標本上,因髓鞘中的類脂被溶解,僅見殘存的蛋白質呈網狀,稱神經角演網(neurokeratin network)。在鋨酸浸染標本上,髓鞘呈黑色,其中還可見數個呈漏斗形的斜裂,稱髓鞘切跡(incisure of myelin)或施-蘭切跡(Schmidt-Lanterman incisure)。電鏡下,髓鞘為明暗相間的同心圓板層排列。髓鞘有保護和絕緣作用,可防 止神經衝動的擴散。
有髓神經纖維的神經衝動傳導,是從一個郎飛結跳到相鄰郎飛結的跳躍式傳導,長的神經纖維,軸突就粗,髓鞘亦厚。結間體也長,傳導速度快。反之,傳導速度慢。大部分腦、脊神經屬於有髓神經纖維。
中樞神經系統有髓神經纖維的髓鞘由少突膠質細胞形成。一個少突膠質細胞的幾個突起,可分別包卷幾條軸突形成髓鞘,其郎飛結較寬,無髓鞘切跡,其胞體位於神經纖維之間。
分類介紹
(1)周圍神經系統的有髓神經纖維:
 神經纖維
神經纖維施萬細胞的胞核呈長卵圓形,其長軸與軸突平行,核周有少量胞質。由於施萬細胞包在軸突的外面,故又稱神經膜細胞(neurilemmal cell),它的外面包有一層基膜。施萬細胞最外面的一層胞膜與基膜一起往往又稱神經膜(neurilemma),光鏡下可見此膜。
髓鞘的形成:在有髓神經纖維發生中,伴隨軸突一起生長的施萬細胞表面凹陷成一縱溝,軸突位於縱溝內,溝緣的胞膜相貼形成軸突系膜(mesaxon)。軸突系膜不斷伸長並反覆包捲軸突,把胞質擠至細胞的內、外邊緣及兩端(即靠近郎氏結處),從而形成許多同心圓的螺鏇膜板層,即為髓鞘。故髓鞘乃成自施萬細胞的胞膜,屬施萬細胞的一部分。施萬細胞的胞質除見於細胞的外、內邊緣和兩端外,還見於髓鞘板層內的施-蘭切跡。該切跡構成螺鏇形的胞質通道,並與細胞外、內邊緣的胞質相通。
(2)中樞神經系統的有髓神經纖維:
其結構基本與周圍神經系統的有髓神經纖維相同,不同的是它的髓鞘不是施萬細胞,而是由少突膠質細胞突起末端的扁平薄膜包捲軸突而形成。一個少突膠質細胞有多個突起可分別包卷多個軸突,其胞體位於神經纖維之間。其次是中樞有髓神經的外表面沒有基膜包裹,髓鞘內亦無施-蘭切跡。
有髓神經的軸膜興奮是呈跳躍式傳導的,故傳導速度快。
有髓神經纖維的髓鞘,因含高濃度類脂而具嫌水性,它不容帶離子的水溶液通過而起絕緣作用。有髓神經纖維軸突的軸膜,除軸突起始段和軸突終末外,只有在郎氏結處才暴露於細胞外環境。其餘大部分的軸膜均被髓鞘包裹。由於髓鞘的電阻比軸膜高得多,而電容卻很低,通過軸突的電流只能使郎氏結處的軸膜去極化而產生興奮。所以,從軸突起始段產生的神經衝動(動作電位)的傳導,是通過郎氏結處的軸膜進行的,即從一個郎氏結跳到下一個郎氏結,呈快速的跳躍式傳導。故而,結間體越長,跳躍的距離也越大,傳導速度也就越快。
無髓神經纖維
無髓神經纖維(nonmyelinated nervefiber)較細的軸突及施萬細胞構成,無髓鞘、無郎飛結。電鏡下可見一個施萬細胞深淺不同的包裹5~15條粗細不等的軸突。無髓神經纖維的神經衝動傳導是沿著軸突進行連續性傳導,其傳導速度比有髓神經纖維慢得多。植物神經的節後纖維和部分感覺神經纖維屬無髓神經纖維。
神經纖維介紹
神經纖維(nerve fiber)是以神經細胞的突起(包括軸突與樹突)為中軸,外包神經膠質細胞(施萬細胞或少突膠質細胞)。根據神經纖維有無髓鞘包裹,分為有髓和無髓神經纖維兩種。
潰變與再生
潰變
神經纖維受損傷如神經被切斷後,切斷處遠側段的神經纖維全長發生潰變(degeneration),軸突和髓鞘碎裂和溶解。與胞體相連的近側段則發生逆行性潰變,即軸突和髓鞘的斷裂溶解由切斷處向胞體方向進行,潰變一般只發展到鄰近斷端的第一側支終止。神經元的胞體腫脹,細胞核從中央移到胞體邊緣,胞質內尼氏體明顯減少,故胞質著色淺淡。
再生
神經元胞體是細胞的營養中心,只有在胞體沒有死亡的條件下才有纖維再生(regeneration)的可能。胞體約於損傷後第3周開始恢復,胞質內的尼氏體重新出現,胞體腫脹消失,胞核恢復中央位置。胞體的完全恢復約需3~6個月,恢復中的胞體不斷合成新的蛋白質及其他產物輸向軸突,使殘留的近側段軸突末端生長出許多新生的軸突支芽。
1.周圍神經纖維的再生 切斷處遠側段的周圍神經纖維,雖然其軸突和髓鞘發生潰變,但包裹神經纖維的基膜仍保留呈管狀。此時施萬細胞大量增生,一面吞噬解體的軸突和髓鞘,一面在基膜管內排列成細胞索,靠近斷口處的施萬細胞還形成細胞橋把兩斷端連線起來。從近側段神經纖維軸突末端長出的軸突支芽,越過此施萬細胞橋,進入基膜管內,當其中一支沿著施萬細胞索生長併到達原來神經纖維末梢所在處,則再生成功。施萬細胞和基膜對軸突的再生起重要的誘導作用。
2.中樞神經纖維的再生 中樞神經纖維的再生比周圍神經困難。神經纖維無施萬細胞,亦無基膜包裹。當中樞神經纖維受損傷時,星形膠質細胞增生肥大,在損傷區形成緻密的膠質瘢痕,大多數再生軸突支不能越過此膠質瘢痕;即使能越過,也沒有如同周圍神經纖維那樣的基膜管和施萬細胞索引導再生軸突到達目的地。所以,中樞神經纖維的損傷常導致脊髓或腦功能的永久性喪失。數十年來不少科學家為研究神經再生進行不懈的努力,近年來已注意到一類能促進神經生長的化學物質稱神經營養因子(neurotrophic factor)的作用。同時又根據胚胎神經元容易生長的化學物質稱神經營養因子(neurotrophic factor)的作用。同時又根據胚胎神經元容易生長及周圍神經能再生的特點,把胚胎腦組織、周圍神經或周圍神經的組分(如基膜或基膜的化學成分)移植到腦內,以期促進中樞神經再生。
在傳導上
在神經細胞中,部分細胞是沒有髓鞘的神經細胞。而在比較高級的動物的神經系統中,神經細胞的軸突部分是由髓鞘包裹著的。而髓鞘並不是完全包裹著軸突,它們是分節的。每一節大約有1mm長,節與節之間有一小段是裸露的部分,這些裸露的部分就被稱為郎飛氏節。在兩段髓鞘之間是無髓鞘的部分,稱為郎飛氏結,其電阻要比結間小得多。因此,在衝動傳導時,局部電流可由一個郎飛氏節跳躍到鄰近的下一個郎飛氏節。這種傳導方式稱為跳躍傳導。跳躍傳導方式極大地加快了傳導的速度。
