代表種
萵苣壞死黃化病毒(Lettucenecroticyellowvirus)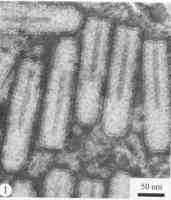 分枝花椰菜壞死黃化病毒
分枝花椰菜壞死黃化病毒病毒特徵
形態學病毒粒子在經過固定後大多為桿菌狀,長200~350nm,直徑70~95nm,未固定樣品呈彈狀或多形性。病毒有包膜,表面具有長5~10nm、直徑約3nm的釘狀突起,它們是由病毒糖蛋白的三
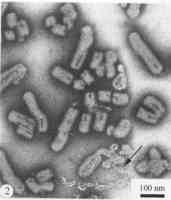 萵苣壞死黃化病毒
萵苣壞死黃化病毒病毒
粗提純的分枝花椰菜壞死黃化病毒(BNYV),中性鉬酸銨負染色。提純的萵苣壞死黃化病毒(LNYV)經磷鎢酸負染色,示部分彈狀粒子結構被破壞,核衣殼從破裂的粒子中流出(箭頭)。理化特性
標準沉降常數S(20W)=940S,在蔗糖中的浮力密度為1.19~1.20g/cm^3。核酸
為單分子線形負義ssRNA,長11~15kb,相對分子質量為4.2×10^6~4.6×10^6。核酸占病毒粒子重量的1%。蛋白質
約占病毒粒子重量的70%,萵苣壞死黃化病毒有5種結構蛋白,分別為蛋白L(分子質量160~180kDa)、蛋白G(78kDa)、蛋白N(57kDa)、蛋白NS(38kDa)和蛋白M(19kDa)。脂類
約占病毒粒子重量的25%,包含在脂蛋白包膜中。碳水化合物
約占病毒粒子重量的3%,與表面突起及糖脂結合。基因組
病毒為單分體基因組,在受侵染細胞的細胞質中複製,與纖維樣結構的病毒基質有關,病毒的形態發生與內質網池有關,在病毒純化時易檢測到內源的轉錄酶活性。該屬只有萵苣壞死黃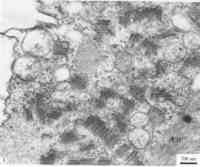 感染萵苣壞死黃化病毒的克利夫蘭菸葉肉細胞
感染萵苣壞死黃化病毒的克利夫蘭菸葉肉細胞抗原特性
免疫原性較弱。已製備出多克隆抗體,針對蛋白G和蛋白N的單克隆抗體用於斑點免疫和ELISA檢測。細胞病理
病毒存在於寄主植物的葉肉、表皮、未成熟感染萵苣壞死黃化病毒的克利夫蘭菸葉肉細胞木質部及韌皮部組織中。感病細胞質內有發達的內質網系統,病毒從內質網膜芽生,成熟的病毒粒子積累在擴大的內質網池中,形成病毒粒子聚集體。病毒不在細胞核內或核周腔中,但有的病毒如萵苣壞死黃化病毒在感染早期細胞核發生變化,核外膜處產生許多小泡結構,核和線粒體、葉綠體等在感染後期有瓦解跡象。有些病毒在細胞質中產生由細顆粒狀或纖維狀物質組成的病毒基質,功能不詳。無外殼或有外殼的粒子也在介體昆蟲的肌肉、脂肪體、菌胞體、氣管、表皮、唾液腺和消化道細胞中觀察到。圖示:感染萵苣壞死黃化病毒的克利夫蘭菸葉肉細胞,示大量病毒粒子處在細胞質膨大的內質網池中,葉綠體和線粒體等細胞器結構開始瓦解。CH,葉綠體;M,線粒體;V,病毒粒子。生物學
寄主範圍單個病毒的寄主範圍較窄,但整個屬的寄主範圍較廣。萵苣壞死黃化病毒的寄主限於藜科、菊科、豆科、百合科和茄科的一些植物。
傳播
萵苣壞死黃化病毒、草莓皺縮病毒、分枝花椰菜壞死黃化病毒等由蚜蟲傳播,大麥黃條點花葉病毒、北方禾穀花葉病毒等由灰飛虱(Laodelphaxstriatellus)、美洲小麥條點花葉病毒由葉蟬傳播,病毒在植物和介體昆蟲體內能增殖。有些病毒可經汁液傳播。地理分布
該屬有的病毒分布是世界性的,有的分布有局限性,萵苣壞死黃化病毒在澳大利亞、紐西蘭有報導。分類
該屬有8個種,代表種是萵苣壞死黃化病毒。中國已報導北方禾穀花葉病毒、草莓皺縮病毒等。編號中文名學名國際基因庫登錄號碼1大麥黃條點花葉病毒Barleyyellowstriatemosaicvirus(BYSMV)-
2分枝花椰菜壞死黃化病毒Broccolinecroticyellowvirus(BNYV)-
3羊茅葉線條病毒Festucaleafstreakvirus(FLSV)-
4萵苣壞死黃化病毒Lettucenecroticyellowsvirus(LNYV)[L24364,L24365,
L30103]
5北方禾穀花葉病毒Northerncerealmosaicvirus(NCMV)
6苦苣菜病毒Sonchusvirus(SonV)
7草莓皺縮病毒Strawberrycrinklevirus(SCV)
8美洲小麥條點花葉病毒WheatAmericanstriatemosaicvirus(WASMV)-
