
定義
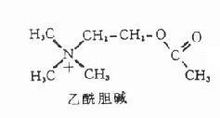 乙醯膽鹼
乙醯膽鹼乙醯膽鹼(acetylcholine,ACh)是一種神經遞質,能特異性的作用於各類膽鹼受體,在組織內迅速被膽鹼酯酶破壞,其作用廣泛,選擇性不高。臨床不作為藥用 。
在神經細胞中,乙醯膽鹼是由膽鹼和乙醯輔酶A在膽鹼乙醯移位酶(膽鹼乙醯化酶)的催化作用下合成的。由於該酶存在於胞漿中,因此乙醯膽鹼在胞漿中合成,合成後由小泡攝取並貯存起來。
進入突觸間隙的乙醯膽鹼作用於突觸後膜發揮生理作用後(乙醯膽鹼可引起受體膜產生動作電位),就被膽鹼酯酶水解成膽鹼和乙酸,這樣乙醯膽鹼就被破壞而推動了作用(迅速分解是為了避免受體細胞膜持續去極化而造成的傳導阻滯),這一過程稱為失活。去甲腎上腺素進入突觸間隙並發揮生理作用後,一部分被血液循環帶走,再在肝中被破壞失活;另一部分在效應細胞內由兒茶酚胺內由兒茶酚胺氧位甲基移位酶和單胺氧化酶的作用而被破壞失活;但大部分是由突觸前膜將去甲腎上腺素再攝取,回收到突觸前膜處的軸漿內並重新加以利用 。
引起乙醯膽鹼量子性釋放的關鍵因素是神經末梢去極化引起的ca2+內流。當神經衝動傳至神經終板時,膜電位下降,導致可使Ca2+ 通過的電壓閘門通道開放,使Ca2+ 進入終板,從而刺激終板分泌乙醯膽鹼。乙醯膽鹼再進一步作用於肌細胞導致肌細胞收縮。
認識
1914年,Ewins在麥角菌中發現了乙醯膽鹼,這是首次在非神經細胞中發現乙醯膽鹼的報導。隨後,人們陸續在多種細菌、真菌、低等植物和高等植物中發現了乙醯膽鹼及其相關的酶和受體。隨著膽鹼能系統在植物中的發現和研究的深入,人們似乎有望在分子水平發現動植物間的又一相似性,因而植物學家抱著極大的熱情投入了這方面的研究。但是由於當時研究手段的限制、對動植物之間的差別認識不足,以及某些研究在其它的實驗室難以重複的緣故,使得植物乙醯膽鹼的研究多處於零星的、非系統的狀態,研究的深度和廣度遠遠無法與動物相比。到目前為止,尚未對其在植物中的作用機理提出一個合理的解釋。 近年來,我們和國外其它幾家實驗室重新開展了乙醯膽鹼在植物體內的生理作用和作用機理的研究,為揭示植物乙醯膽鹼的作用機理提供了新的線索。
植物生理過程
【對代謝、生長和發育的調控】
種子萌發
 乙醯膽鹼
乙醯膽鹼生長
乙醯膽鹼對於生長的影響因實驗條件的不同,植物種類或同一植物不同組織而異。乙醯膽鹼可以模擬紅光的作用抑制大豆側根的發育,還可以引起小麥幼苗生長和乾重的增加。在離體組織中,乙醯膽鹼可以刺激燕麥胚芽鞘和黃瓜下胚軸的伸長以及綠豆下胚軸的生長,刺激蠶豆下胚軸的生長而抑制其上胚軸的生長。總之,乙醯膽鹼對植物生理過程的影響與所利用的組織及實驗條件密切相關,其最大效應在pH酸性區。
成花作用
乙醯膽鹼可以模擬紅光的作用,抑制遠紅光刺激的過氧化物酶活性升高,從而使菠菜能在非誘導條件下開花。乙醯膽鹼可以抑制連續光照條件下(24 h光照/0 h黑暗)長日浮萍G1的成花和刺激非誘導的短日條件下短日浮萍Torr的成花。阿托品可以抑制連續光照下生長的青萍G3成花而管箭毒則無作用,說明乙醯膽鹼對成花的誘導作用可能是通過質膜上的類毒蕈鹼型受體介導的。乙醯膽鹼對成花的誘導作用還可能與它調控的膜對離子的通透性有關。光周期誘導的成花作用涉及到葉片膜電勢的改變,乙醯膽鹼還可能通過影響膜電勢而參與成花誘導。
呼吸作用
乙醯膽鹼可引起根尖細胞耗氧速率的增加。Jaffe以游離的線粒體為材料得到的結果已證實了這一點。伴隨著氧的消耗,組織中ATP的水平下降10倍,自由磷水平升高14倍。乙醯膽鹼的這種作用可能是其使呼吸的電子傳遞鏈與氧化磷酸化解偶聯所造成。根據這些實驗結果,Jaffe提出了乙醯膽鹼對大豆根尖細胞的作用模式,即當胞間乙醯膽鹼濃度升高時,乙醯膽鹼到達其作用的靶部位,隨後是分泌質子,氧的消耗和ATP水解增加,而這些過程均與膜對陽離子通透性的增加相關連。
光合作用
乙醯膽鹼可以在不影響電子傳遞的情況下使葉綠體中的ATP合成下降80%以上。另外,濃度低於0.1 mmol的乙醯膽鹼可以刺激非環式光合磷酸化的進行,而濃度大於0.1 mmol時非環式光合磷酸化則受抑制。在這兩種情況下,乙醯膽鹼並不影響NADP+的還原。新斯的明(neostigmine)可以抑制ATP的合成,但不影響電子從水到細胞色素f或NADP+的傳遞。毒蕈鹼和阿托品同樣可以抑制NADP+的還原和非環式光合磷酸化。
除此以外,乙醯膽鹼還可以影響離體葉綠體對氧的吸收,抑制光刺激的葉綠體膨脹;刺激鈉離子和鉀離子從葉綠體流出。因而,乙醯膽鹼在葉綠體中可能調控葉綠體膜對離子的通透性及電子傳遞和ATP合成間的偶聯。
【對與膜透性有關的生理過程的調控】
棚田效應
紅光促使黃化的綠豆和大麥根尖吸附到帶負電的玻璃杯內壁上,而遠紅光則使根尖脫離杯壁釋放到溶液中。這種現象稱為棚田效應(tanada effect)。 在黑暗中乙醯膽鹼可以使離體的大豆根尖吸附到帶負電的玻璃杯內壁上,並阻止遠紅光引起的根尖脫離杯壁,乙醯膽鹼酯酶抑制劑毒扁豆鹼(eserine)增加組織對乙醯膽鹼的敏感性。這些說明內源乙醯膽鹼可能在這一生理過程中有控制作用。
紅光可提高組織中乙醯膽鹼水平,其原因可能與紅光促進Pfr的形成有關,而後者則與乙醯膽鹼合成有關。組織中乙醯膽鹼水平升高可以刺激質子從根細胞流出到溶液中,從而形成表面正電勢,以致根尖被吸附到帶負電的玻璃杯內壁上;遠紅光促使光敏素從遠紅光吸收型(Pfr)轉變為紅光吸收型(Pr),致使根尖從玻璃杯內壁釋放到溶液中。但是也有實驗指出乙醯膽鹼在這一過程中僅相當於單價陽離子的作用。
葉片運動
Jaffe提出乙醯膽鹼可能調控含羞草葉片的運動。紫花大翼豆是一種常用的牧草,在強光照下其葉片可以下垂以避免高光強對葉片的直接傷害。據報導,強光下來源於熱帶的品種比來自溫帶品種的葉片下垂快,光強減弱後下垂狀態恢復更快。測定此種植物葉褥組織中乙醯膽鹼的結果表明,乙醯膽鹼水平的變化與葉片的狀態密切相關。來源於熱帶品種的含量和變化幅度更大,外施乙醯膽鹼可以使其從下垂狀態中恢復。進一步研究的結果顯示,葉片中乙醯膽鹼水平的變化是由乙醯膽鹼酯酶控制的,而乙醯膽鹼酯酶主要分布於維管束的周圍,因而推測乙醯膽鹼可能通過影響離子進出維管束,進而影響水分的進出最終實現葉片運動的調控。
通透性
乙醯膽鹼可以刺激質子從大豆根尖細胞流出,誘導菠菜葉片膜電勢的變化,抑制藍光誘導的大豆下胚軸彎鉤膜電勢的超極化及該組織對鉀的吸收,這些過程都涉及乙醯膽鹼對膜透性的調節。
除了影響上述過程外,乙醯膽鹼還可以影響組織對鈣離子的吸收。Tretyn發現乙醯膽鹼可以刺激黃化燕麥胚芽鞘對鈣離子的吸收。乙醯膽鹼酯酶的抑制劑可以增加組織對乙醯膽鹼的敏感性;鈣通道的抑制劑可以抑制乙醯膽鹼刺激的鈣吸收的增加。這些結果表明乙醯膽鹼參與調控植物的鈣通道。
代謝的影響
乙醯膽鹼可以影響植物的膜脂代謝。如它可以抑制磷摻入到黃化大豆莖切段的磷脂分子中,但在有氧條件下主要抑制磷摻入磷脂醯乙醇胺和磷脂醯膽鹼,而在無氧條件下乙醯膽鹼主要抑制磷摻入磷脂醯肌醇。這些結果表明植物的磷脂與動物的磷脂間有相似性,乙醯膽鹼可以同樣影響植物的磷脂代謝。
參與相互作用
參與植物與植物以及細胞與細胞之間的相互作用
在一個生態環境中,植物與植物之間以及植物與其他生物之間常常表現出相互作用的關係。這種相互作用可以是促進性的也可以是抑制性的,即表現為相生相剋的關係。乙醯膽鹼酯酶存在於根瘤菌感染大豆所形成的根瘤中,而且乙醯膽鹼酯酶的最大活性與根瘤對氮的最大同化期相一致,推測乙醯膽鹼及其酯酶在根瘤菌和寄主植物間的相互作用中起一定作用。乙醯膽鹼酯酶還存在於地衣的葉狀體中,而且主要分布於組成地衣的真菌和藻類兩種生物的界面。在其粉芽(soredia)產生孢子過程中,乙醯膽鹼酯酶活性增加,而且酶活性集中分布在接觸區。乙醯膽鹼及乙醯膽鹼酯酶參與地衣這種生物間相互作用的機理可能是通過調控膜對離子的通透性,並介導環境中光對地衣生殖影響而實現的乙醯膽鹼及乙醯膽鹼酯酶還可能參與花粉與柱頭間的識別作用。在裂葉牽牛中乙醯膽鹼酯酶主要分布在雌蕊柱頭的表面,還存在於花粉粒和花粉管的尖端。乙醯膽鹼的激活劑和拮抗劑以及乙醯膽鹼酯酶的抑制劑均可以影響某些植物的花粉萌發和花粉管伸長。因此推測乙醯膽鹼和乙醯膽鹼酯酶可能參與植物柱頭和花粉間的相互作用。
作用機理
在動物細胞中乙醯膽鹼與受體結合後,一方面直接影響膜對離子的通透性,另一方面通過各種第二信使影響各種生理過程的進行。在植物界中,雖然乙醯膽鹼的受體還沒有在生化上最後確定,但是一系列藥理學的證據表明植物中確實存在著乙醯膽鹼的受體。關於植物中乙醯膽鹼與受體結合後的事件了解甚少。有關乙醯膽鹼在植物細胞中作用機理的研究,除上面的零散研究外,只有Tretyn等在小麥原生質體膨脹和幼葉展開中的研究較為系統,並揭示了其中的某些細節。
信號轉導
原生質體膨脹,紅光可以刺激黃化小麥葉肉細胞原生質體體積膨脹,這種刺激作用可為隨後的遠紅光照射所逆轉,說明這一反應是在光敏素控制下進行的。紅光對原生質體體積膨脹的刺激作用要求介質中含有Ca2+。乙醯膽鹼可以代替紅光在黑暗中引起原生質體的膨脹。與紅光引起的反應不同,乙醯膽鹼不僅可以在含Ca2+的介質中引起原生質體的膨脹,而且在含Na+或K+的介質中也可以引起原生質體的膨脹。
除乙醯膽鹼外,只有氨基甲醯膽鹼可以刺激原生質體的膨脹,而膽鹼、丙醯膽鹼和丁醯膽鹼則無此作用。乙醯膽鹼酯酶的抑制劑毒扁豆鹼可以增加原生質體對乙醯膽鹼的敏感性。據此可以認為乙醯膽鹼能特異地刺激黃化小麥葉肉原生質體膨脹
乙醯膽鹼誘導原生質體膨脹過程中是否涉及乙醯膽鹼受體的參與可用乙醯膽鹼受體的激活劑和抑制劑來確定。乙醯膽鹼N型受體的激活劑菸鹼在含Na+或K+的介質中可以直接刺激原生質體膨脹,而在含Ca2+的介質中,菸鹼沒有作用。與以上結果不同,M型受體的激活劑毒蕈鹼可以在含Ca2+的介質中刺激原生質體膨脹,而在含Na+或K+的介質中沒有作用。採用乙醯膽鹼受體抑制劑的研究也得出同樣的結論。M型受體的抑制劑阿托品在含Na+或K+的介質中對乙醯膽鹼刺激的原生質體膨脹沒有作用,但在含Ca2+介質中則可以抑制乙醯膽鹼誘導的原生質體膨脹。N型受體抑制劑管箭毒在含Ca2+介質中對乙醯膽鹼刺激的原生質體膨脹沒有作用,但在含Na+或K+的介質中則有抑制作用。螢光定位技術證明N型乙醯膽鹼受體主要分布在原生質體表面。
在乙醯膽鹼誘導的原生質體膨脹過程中,乙醯膽鹼為受體接受後的信號轉導可能涉及到Ca2+ 和CaM,因為Ca2+ 通道抑制劑尼群地平(nifedipine, NIF)和La3+ 可以完全抑制乙醯膽鹼誘導的原生質體在含Ca2+ 介質中的膨脹。同樣,鈣調素的抑制劑和G蛋白的抑制劑也有這樣的作用,而這些化合物在含Na+ 或K+ 的介質中則沒有作用。
幼葉展開,生長於黑暗中8 d的小麥幼苗,其初生葉的展開受控於光敏色素系統。如果介質中含有Ca2+ ,乙醯膽鹼在暗中可以刺激離體葉切段中幼葉的展開。在沒有Ca2+ 而有Na+ 的介質中乙醯膽鹼也可以刺激黃化小麥初生葉片的展開。在乙醯膽鹼的各種衍生物中只有氨基甲醯乙醯膽鹼可以刺激黃化小麥初生葉片的展開。乙醯膽鹼受體的拮抗劑,阿托品和D-管箭毒可以分別抵消乙醯膽鹼在含Ca2+ 和Na+ 介質中誘導葉片的展開。乙醯膽鹼受體的激活劑,毒蕈鹼和菸鹼可以分別在Ca2+ 和Na+ 的介質中刺激原生質體的膨脹。乙醯膽鹼誘導的Ca2+ 依賴的葉片開展可為Ca2+ 通道抑制劑尼群地平和鈣調素抑制劑3-氟-甲基吩噻嗪(trifluoperazine, TFP)所減弱,其中只有鈣調素抑制劑TFP可以抑制乙醯膽鹼誘導的在含Na+ 介質中黃化小麥初生葉片的展開。
根據以上結果可以初步認為,在植物中乙醯膽鹼可能以一種類似於在動物中的機制發揮作用。乙醯膽鹼首先與M型和N型乙醯膽鹼受體結合。與在動物細胞中一樣,M型乙醯膽鹼受體可能與磷酸肌醇代謝途徑有關,在此途徑中,G蛋白,Ca2+ 通道和鈣調素等相繼被激活,最後發生生理反應。N型受體是非磷酸肌醇依賴的,它直接控制膜對離子的通透性。這兩條途徑可以相互獨立地引起原生質體的膨脹或葉片的張開。
酶活性
乙醯膽鹼在植物中的作用機理除參與調節膜對離子的通透性外,可能還涉及對植物體內某些酶活性的調控。乙醯膽鹼對兵豆(Lens culinaris)根生長的抑制作用與體內過氧化物同工酶的活性變化密切相關,它可以刺激某些同工酶的活性而抑制另外一些同工酶的活性。
乙醯膽鹼本身對於植物體內苯丙氨酸氨基裂解酶的活性和類黃酮的合成沒有影響,但它卻可以抵消紅光對苯丙氨酸氨基裂解酶活性和類黃酮合成的刺激作用。
【對內源生長調節物質的影響】
乙醯膽鹼可以影響植物體內吲哚乙酸和乙烯的代謝。在大豆下胚軸中,乙醯膽鹼抑制吲哚乙酸刺激的乙烯合成並抵消它對大豆下胚軸彎鉤伸直的抑制作用,它也可以抵消乙烯刺激的蕨類植物原絲體的生長。乙醯膽鹼的這種作用可能是通過影響內源吲哚乙酸和乙烯的水平而實現的。以離體大豆葉片的實驗證明乙醯膽鹼可以抑制組織中乙烯的合成。
乙醯膽鹼還可能與內源的赤黴素相互作用。它可以部分代替赤黴素誘導黃瓜下胚軸的伸長,還可以引起植物體內游離態的赤黴素含量增高,這種增高可以阿托品抵消。
生理活動影響
中樞膽鹼能系統與學習、記憶密切相關,乙醯膽鹼(ACh)是中樞膽鹼能系統中重要的神經遞質之一,其主要功能是維持意識的清醒,在學習記憶中起重要作用。
認知活動關係
腦內細胞外乙醯膽鹼(ach)的變化主要反映膽鹼能神經元的活動,皮層和海馬等腦區的ach主要來源於基底前腦膽鹼能神經元的纖維投射。套用微透析等技術在體檢測清醒、自由活動動物認知過程中腦內乙醯膽鹼的含量,可以研究ach與特定行為反應和認知活動之間的關係。研究發現當機體需要對新刺激進行分析時,在學習與記憶、空間工作記憶、注意、自發運動和探究行為等認知活動中,基底前腦膽鹼能神經元被激活,腦內ach的釋放也隨之改變。結果提示腦內膽鹼能遞質系統活動與認知過程密切相關。
帕金森病關係
帕金森病又稱"震顫麻痹",是一種中樞神經系統變性疾病,主要是因位於中腦部位"黑質"中的細胞發生病理性改變後,多巴胺的合成減少,抑制乙醯膽鹼的功能降低,則乙醯膽鹼的興奮作用相對增強。兩者失衡的結果便出現了"震顫麻痹"。
黑質細胞發生變性壞死的原因迄今尚未明了, 可能與遺傳和環境因素有關。有學者認為蛋白質、水果、乳製品等攝入不足,嗜酒、外傷、過度勞累及某些精神因素等,均可能是致病的危險因素。 原因不明的多巴胺減少導致的震顫麻痹,在醫學上稱為"原發性震顫麻痹",即帕金森病;
人體利用
人的腦組織有大量乙醯膽鹼,但乙醯膽鹼的含量會隨著年齡的增加出下降。正常老人比青年時下降30%,而老年痴呆患者下降更為嚴重,可達70%~80%。美國醫生伍特曼觀察到老年人腦組織乙醯膽鹼減少,就給老年人吃富含膽鹼的食品,發現有明顯的防止記憶減退的作用。英國和加拿大等國的科學家也相繼進行了研究,一致認為只要有控制地供給足夠的膽鹼,可避免60歲左右老年人記憶力減退。所以保持和提高大腦中乙醯膽鹼的含量,是解決記憶力下降的根本途徑。在自然界是,乙醯膽鹼多以膽鹼的狀態存在於蛋、魚、肉、大豆等之中,這些膽鹼必須在人體內起生化反應後,才能合成具有生理活性的乙醯膽鹼。另外,經常服用蜂王漿可以提高腦內乙醯膽鹼的含量,從而促進激活腦神經傳導功能,提高信息傳遞速度,增強大腦記憶能力,全面改善腦功能,並能延緩衰老。
藥理作用
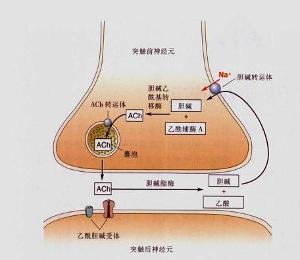 乙醯膽鹼在突觸的合成與作用
乙醯膽鹼在突觸的合成與作用1.心血管系統ACh對心血管系統主要產生以下作用:
(1)血管擴張作用:靜注小劑量本品可由於全身血管擴張而造成血壓短暫下降,並伴有反射性心率加快。ACh可引起許多血管擴張。如肺和冠狀血管。其擴血管作用主要由於激動血管內皮細胞M,膽鹼受體亞型,導致內皮依賴性舒張因子(EDRF)即一氧化氮(nitric oxide,No)釋放,從而引起鄰近平滑肌細胞鬆弛,也可能通過壓力感受器或化學感受器反射引起。如果血管內皮受損,則ACh的上述作用將不復存在,相反可引起血管收縮。此外,ACh通過激動交感神經末梢突觸前膜M1受體,抑制去甲腎上腺素能神經末梢釋放NA也與ACh擴血管作用有關。
(2)減慢心率:亦稱負性頻率作用。ACh能使竇房結舒張期自動除極延緩,復極化電流增加,使動作電位到達閾值的時間延長,導致心率減慢。
(3)減慢房室結和普肯耶纖維傳導:即為負性傳導作用。ACh可延長房室結和普肯耶纖維(Βurkinje fibers)的不應期,使其傳導減慢。當使用強心苷使迷走神經張力增高或全身給藥法使用大劑量膽鹼受體激動藥時所出現的完全性心臟傳導阻滯常與房室結傳導明顯抑制有關。
(4)減弱心肌收縮力:即為負性肌力作用。一般認為膽鹼能神經主要分布於竇房結、房室結、普肯耶纖維和心房,而心室較少有膽鹼能神經支配,故ACh對心房收縮的抑制作用大於心室。但由於迷走神經末梢與交感神經末梢緊密相鄰,迷走神經末梢所釋放的ACh可激動交感神經末梢突觸前M膽鹼受體,反饋性抑制交感神經末梢去甲腎上腺素釋放。使心室收縮力減弱。
(5)縮短心房不應期:ACh不影響心房肌的傳導速度,但可使心房不應期及動作電位時程縮短(即為迷走神經作用)。
2.胃腸道
ACh可明顯興奮胃腸道平滑肌,使其收縮幅度、張力、蠕動增加,並可促進胃、腸分泌,引起噁心、暖氣、嘔吐、腹痛及排便等症狀。
3.泌尿道
ACh可使泌尿道平滑肌蠕動增加,膀胱逼尿肌收縮,使膀胱最大自主排空壓力增加,降低膀胱容積,同時膀胱三角區和外括約肌舒張,導致膀胱排空。
4.其他
(1)腺體:ACh可使淚腺、氣管和支氣管腺體、唾液腺、消化道腺體和汗腺分泌增加。
(2)眼:當ACh局部滴眼時,可致瞳孔收縮,調節於近視。
(3)神經節和骨骼肌:ACh作用於自主神經節NN膽鹼受體和骨骼肌神經肌肉接頭的NM膽鹼受體,引起交感、副交感神經節興奮及骨骼肌收縮。此外,因腎上腺髓質受交感神經節後纖維支配,故NN膽鹼受體激動能引起腎上腺索釋放。
(4)中樞:由於ACh不易進人中樞,故儘管中樞神經系統有膽鹼受體存在,但外周給藥很少產生中樞作用。
(5)支氣管:ACh可使支氣管收縮。
(6)ACh還能興奮頸動脈體和主動脈體化學受體。
盤點盜夢空間的背景知識
| 本季扛鼎大片《盜夢空間》在今天正式上映了,晦澀難懂撲朔迷離的劇情猶如第二部《黑客帝國》。 |
