
概述
 骨骼肌
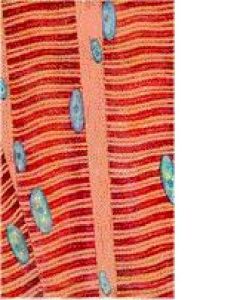
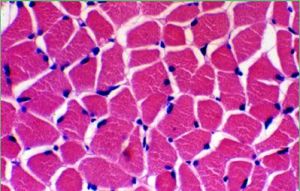
骨骼肌相鄰的各肌原纖維,明帶均在一個平面上,暗帶也在一個平面上,因而使肌纖維顯出明暗相間的橫紋。骨骼肌細胞構成骨胳肌組織,每塊骨骼肌主要由骨骼肌組織構成,外包結締組織膜、內有神經血管分布。骨骼肌收縮受意識支配,故又稱“隨意肌”。收縮的特點是快而有力,但不持久。
運動系統的肌肉muscle屬於橫紋肌,由於絕大部分附著於骨,故又名骨骼肌。每塊肌肉都是具有一定形態、結構和功能的器官,有豐富的血管、淋巴分布,在軀體神經支配下收縮或舒張,進行隨意運動。肌肉具有一定的彈性,被拉長後,當拉力解除時可自動恢復到原來的程度。肌肉的彈性可以減緩外力對人體的衝擊。肌肉內還有感受本身體位和狀態的感受器,不斷將衝動傳向中樞,反射性地保持肌肉的緊張度,以維持體姿和保障運動時的協調。
分類
 骨骼肌
骨骼肌分為頭肌、軀幹肌、四肢肌等。
2.根據肌的功能
分為屈肌、伸肌、內收肌、外展肌、鏇內肌、鏇外肌等。
3.根據肌的外形
分為長肌、短肌、扁肌和輪匝肌等。
構造形態
 骨骼肌
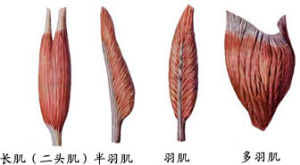
骨骼肌肌的形態各異,有長肌、短肌、闊肌、輪匝肌等基本類型。長肌多見於四肢,主要為梭形或扁帶狀,肌束的排列與肌的長軸相一致,收縮的幅度大,可產生大幅度的運動,但由於其橫截面肌束的數目相對較少,故收縮力也較小;另有一些 肌有長的腱,肌束斜行排列於腱的兩側,酷似羽毛名為羽狀肌(如股直肌),或斜行排列於腱的一側,叫半羽狀肌(如半膜肌、拇長屈肌),這些肌肉其生理橫斷面肌束的數量大大超過梭形或帶形肌,故收縮力較大,但由於肌束短,所以運動的幅度小。短肌多見於手、足和椎間。闊肌多位於軀幹,組成體腔的壁。輪匝肌則圍繞於眼、口等開口部位。
命名原則
肌肉可根據共形狀、大小、位置、起止點、纖維方向和作用等命名。依形態命名的如斜方肌、菱形肌、三角肌、梨狀肌等;依位置命名的如肩胛下肌、岡上肌、岡下肌、肱肌等;依位置和大小綜合命名的有胸大肌、胸小肌、臀大肌等;依起止點命名的如胸鎖乳突肌、肩胛舌骨肌等;依纖維方向和部位綜合命名的有腹外斜肌、肋間外肌等;依作用命名的如鏇後肌、咬肌等;依作用結合其它因素綜合命名的如鏇前圓肌、內收長肌、指淺屈肌等。結構關係
 骨骼肌
骨骼肌一般講幾軸性關節就具有與幾個運動軸相對應的對抗肌群,但也有個別關節,有的運動軸沒有相應肌肉配布,如手的掌指關節,從關節面的形態看屬於球窩關節,卻只生有屈伸和收展兩組對抗的肌肉,而沒有與垂直軸交叉的迴旋肌,所以該關節不能做主動的迴旋運動,當然它有一定的被動的迴旋能力。
圍繞某一個運動軸作用相反的兩組肌肉叫做對抗肌,但在進行某一運動時,一組肌肉收縮的同時,與其對抗的肌群則適度放鬆並維持一定的緊張度,二者對立統一,相反相成。另外,在完成一個運動時,除了主要的運動肌(原動肌)收縮外,尚需其它肌肉配合共同完成,這些配合原動肌的肌肉叫協力肌。當然,肌肉彼此間的關係,往往由於運動軸的不同,它們之間的關係也是互相轉化的,在沿此一軸線運動時的兩個對抗肌,到沿彼一軸線運動時則轉化為協力肌。
此外,還有一些運動,在原動肌收縮時,必須另一些肌肉固定附近的關節,如握緊拳的動作,需要伸腕肌將腕關節固定在伸的位置上,屈指肌才能使手指充分屈曲將拳握緊,這種不直接參與該動作而為該動作提供先決條件的肌肉叫做共濟肌。
光鏡結構
骨骼肌纖維
 骨骼肌纖維
骨骼肌纖維肌原纖維
肌原纖維(myofibril)呈細絲狀,直徑1~2μm,沿肌纖維長軸平行排列,每條肌原纖維上都有明暗相間、重複排列的橫紋(crossstriation)。由於各條肌原纖維的明暗橫紋都相應地排列在同一平面上,因此肌纖維呈現出規則的明暗交替的橫紋。橫紋由明帶和暗帶組成。在偏光顯微鏡下,明帶(lightband)呈單折光,為各向同性(isotropic),又稱I帶;暗帶(darkband)呈雙摺光,為各向異性(anisotropic),又稱A帶。在電鏡下,暗帶中央有一條淺色窄帶稱H帶,H帶中央還有一條深M線。明帶中央則有一條深色的細線稱Z線。兩條相鄰Z線之間的一段肌原纖維稱為肌節(sarcomere)。每個肌節都由1/2I帶+A帶+1/2I帶所組成。肌節長約2~2.5μm,它是骨骼肌收縮的基本結構單位。因此,肌原纖維就是由許多肌節連續排列構成的。
超微結構
肌原纖維
肌原纖維是由上千條粗、細兩種肌絲有規律地平行排列組成的,明、暗帶就是這兩種肌絲排布的結果。粗肌絲(thickfilament)長約1.5μm,直徑約15nm,位於肌節的A帶。粗肌絲中央借M線固定,兩端游離。細肌絲(thinfilathent)長約1μm,直徑約5nm,它的一端固定在Z線上,另一端插入粗肌絲之間,止於H帶外側。因此,I帶內只有細肌絲,A帶中央的H帶內只有粗肌絲,而H帶兩側的A帶內既有粗肌絲又有細肌絲;所以在此處的橫切面上可見一條粗肌絲周圍有6條細肌絲;而一條細肌絲周圍有3條粗肌絲。兩種肌絲肌在肌節內的這種規則排列以及它們的分子結構,是肌纖維收縮功能的主要基礎。
分子結構
粗肌絲
 骨骼肌細胞
骨骼肌細胞M線兩側的肌球蛋白對稱排列,桿部均朝向粗肌絲的中段,頭部則朝向粗肌絲的兩端的兩端並露出表面,稱為橫橋(crossbridge)。M線兩側的粗肌絲只有肌球蛋白桿部而沒有頭部,所以表面光滑。肌球蛋白頭部是一種ATP酶,能與ATP結合。只有當肌球蛋白分子頭部與肌動蛋白接觸時,ATP酶才被激活,於是分解ATP放出能量,使橫橋發生屈伸運動。
細肌絲
細肌絲由三種蛋白質分子組成,即肌動蛋白、原肌球蛋白和肌原蛋白。後二種屬於調節蛋白,在肌收縮中起調節作用。肌動蛋白(actin)分子單體為球形,許多單體相互接連成串珠狀的纖維形,肌動蛋白就是由兩條纖維形肌動蛋白纏繞形成的雙股螺鏇鏈。每個球骨骼肌形肌動蛋白單體上都有一個可以與肌球蛋白頭部相結合的位點。原肌球蛋白(tropomyosin)是由較短的雙股螺鏇多肽鏈組成,首尾相連,嵌於肌動蛋白雙股螺鏇鏈的淺溝內。肌原蛋白(troponin)由3個球形亞單位組成,分別簡稱為TnT、TnI和TnC。肌原蛋白借TnT而附於原肌球蛋白分子上,TnI是抑制肌動蛋白和肌球蛋白相互作用的亞單位,TnC則是能與Ca2+相結合的亞單位。
橫小管
它是肌膜向肌漿內凹陷形成的小管網,由於它的走行方向與肌纖維長軸垂直,故稱橫小管(transversetubule,或稱T小管)。人與哺乳動物的橫小管位於A帶與I帶交界處,同一水平的橫小管在細胞內分支吻合環繞在每條肌原纖維周圍。橫小管可將肌膜的興奮迅速傳到每個肌節。
肌漿網
肌漿網(sarcoplasmicreticulum)是肌纖維內特化的滑面內質網,位於橫小管之間,縱行包繞在每條肌原纖維周圍,故又稱縱小管。位於橫小管兩側的肌漿網呈環行的扁囊,稱終池(terminalcisternae),終池之間則是相互吻合的縱行小管網。每條橫小管與其兩側的終池共同組成骨骼肌三聯體(triad)。在橫小管的肌膜和終池的肌漿網膜之間形成三聯體連線,可將興奮從肌膜傳到肌漿網膜。肌漿網的膜上有豐富的鈣泵(一種ATP酶),有調節肌漿中Ca2+濃度的作用。
輔助結構
筋膜
 骨骼肌
骨骼肌深筋膜又叫固有筋膜,由緻密結締組織構成,遍布全身,包裹肌肉、血管神經束和內臟器官。深筋膜除包被於肌肉的表面外,當肌肉分層時,固有筋膜也分層。在四肢,由於運動較劇烈,固有筋膜特別發達、厚而堅韌,並向內伸入直抵骨膜,形成筋膜鞘將作用不同的肌群分隔開,叫做肌間隔。在體腔肌肉的內面,也襯以固有筋膜,如胸內、腹內和盆內筋膜等,甚而包在一些器官的周圍,構成臟器筋膜。
一些大的血管和神經乾在肌肉間穿行時,深筋膜也包繞它們,形成血管鞘。筋膜的發育與肌肉的發達程度相伴行,肌肉越發達,筋膜的發育也愈好。在手腕及足踝部,固有筋膜增厚形成韌帶並伸入深部分隔成若干隧道,以約束深面通過的肌腱。在筋膜分層的部位,筋膜之間的間隙充以疏鬆結締組織,叫做筋膜間隙,
腱鞘
一些運動劇烈的部位如手和足部,長肌腱通過骨面時,其表面的深筋膜增厚,並伸向深部與骨膜連線,形成筒狀的纖維鞘,其內含由滑膜構成的雙層圓筒狀套管,套管的內層緊包在肌腱的表面,外層則與纖維鞘相貼。兩層之間含有少量滑液。因此肌腱既被固定在一定位置上,又可滑動並減少與骨面的摩擦。在發生中滑膜鞘的兩層在骨面與肌腱間互相移行,叫做腱系膜,發育過程中腱系膜大部分消失,僅在一定部位上保留,以引導營養肌腱的血管通過。滑液囊
在一些肌肉抵止腱和骨面之間,生有結締組織小囊,壁薄,內含滑液,叫做滑液囊。其功能是減緩肌腱與骨面的摩擦。滑液囊有的是獨立封閉的,有的與鄰近的關節腔相通,可視為關節囊滑膜層的突出物。收縮原理
 骨骼肌
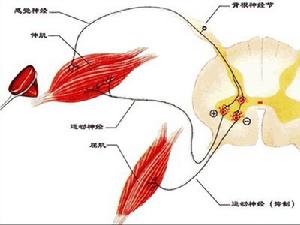
骨骼肌①運動神經末梢將神經衝動傳遞給肌膜;
②肌膜的興奮經橫小管迅速傳向終池;
③肌漿網膜上的鈣泵活動,將大量Ca2+轉運到肌漿內;
④肌原蛋白TnC與Ca2+結合後,發生構型改變,進而使原肌球蛋白位置也隨之變化;
⑤原來被掩蓋的肌動蛋白位點暴露,迅即與肌球蛋白頭接觸;
⑥肌球蛋白頭ATP酶被激活,分解了ATP並釋放能量;
⑦肌球蛋白的頭及桿發生屈曲轉動,將肌動蛋白拉向M線;
⑧細肌絲向A帶內滑入,I帶變窄,A帶長度不變,但H帶因細肌絲的插入可消失,由於細肌絲在粗肌絲之間向M線滑動,肌節縮短,肌纖維收縮;
⑨收縮完畢,肌漿內Ca2+被泵入肌漿網內,肌漿內Ca2+濃度降低,肌原蛋白恢復原來構型,原肌球蛋白恢復原位又掩蓋肌動蛋白位點,肌球蛋白頭與肌動蛋白脫離接觸,肌則處於鬆弛狀態。
運動機理
 骨骼肌
骨骼肌運動神經纖維在到達神經末梢處時先失去髓鞘,以裸露的軸突末梢嵌入到肌細胞膜上稱作終板的膜凹陷中,但軸突末梢的膜和終板膜並不直接接觸,而是被充滿了細胞外液的接頭間隙隔開,其中尚含有成分不明的基質;有時神經末梢下方的終板膜還有規則地再向細胞內凹入,形成許多皺褶,其意義可能在於增加接頭後膜的面積,使它可以容納較多數目的蛋白質分子,它們最初被稱為N-型乙醯膽鹼受體,現已證明它們是一些化學門控通道,具有能與ACh特異性結合的亞單位。在軸突末梢的軸漿中,除了有許多線粒體外還含有大量直徑約50nm的無特殊構造的囊泡。用組織化學的方法可以證明,囊泡內含有ACh;此ACh首先在軸漿中合成,然後貯存在囊泡內。
據測定,每個囊泡中貯存的ACh量通常是相當恆定的,且當它們被釋放時,也是通過出胞作用,以囊泡為單位“傾囊”釋放,被稱為量子式釋放。在神經末梢處於安靜狀態時,一般只有少數囊泡隨機地進行釋放,不能對肌細胞產生顯著影響。但當神經末梢處有神經衝動傳來時,在動作電位造成的局部膜去極化的影響下,大量囊泡向軸突膜的內側面靠近,通過囊泡膜與軸突膜的融合,並在融合處出現裂口,使囊泡中的ACh全部進入接頭間隙。
據推算,一次動作電位的到達,能使大約200~300個囊泡的內容排放,使近107個ACh分子被釋放。軸突末梢處的電位變化引起囊泡排放的過程十分複雜,但首先是軸突末梢膜的去極化,引起了該處特有的電壓門控式Ca2+通道開放,引起細胞間隙液中的Ca2+進入軸突末梢,觸發了囊泡移動以至排放的過程。Ca2+的進入量似乎決定著囊泡釋放的數目;細胞外液中低Ca2+或(和)高Mg2+,都可阻礙ACh的釋放而影響神經-肌接頭的正常功能。已故馮德培院士在30年代對神經-肌接頭的化學性質傳遞進行過重要的研究。
特性
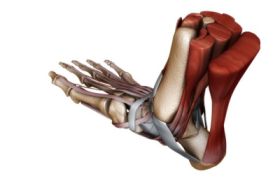 骨骼肌
骨骼肌1、伸展性:骨骼肌在受到外力牽拉或負重時可被拉長的特性。
2、彈性:當外力或負重取消後,肌肉的長度又可恢復的特性。
3、粘滯性:由於肌漿內各分子之間的相互摩擦作用所產生的特性。
生理特性
1、興奮性:骨骼肌是可興奮組織,受到刺激後可產生興奮(即產生動作電位)的特性。
2、收縮性:肌肉受到刺激產生興奮後,立即產生收縮反應的特性引起骨骼肌興奮刺激條件。
3、刺激強度:要使肌肉產生興奮,刺激必須達到一定強度閾上刺激閾下刺激。
4、閾刺激:引起肌肉興奮的最小刺激強度刺激的作用時間刺激強度變化率要使可興奮組織興奮,刺激必須有足夠的變化率。
運動神經
運動神經纖維在到達神經末梢處時先失去髓鞘,以裸露的軸突末梢嵌入到肌細胞膜上稱作終板的膜凹陷中,但軸突末梢的膜和終板膜並不直接接觸,而是被充滿了細胞外液的接頭間隙隔開,其中尚含有成分不明的基質;有時神經末梢下方的終板膜還有規則地再向細胞內凹入,形成許多皺褶,其意義可能在於增加接頭後膜的面積,使它可以容納較多數目的蛋白質分子,它們最初被稱為N-型乙醯膽鹼受體,現已證明它們是一些化學門控通道,具有能與ACh特異性結合的亞單位。在軸突末梢的軸漿中,除了有許多線粒體外還含有大量直徑約50nm的無特殊構造的囊泡。在神經末梢處於安靜狀態時,一般只有少數囊泡隨機地進行釋放,不能對肌細胞產生顯著影響。但當神經末梢處有神經衝動傳來時,在動作電位造成的局部膜去極化的影響下,大量囊泡向軸突膜的內側面靠近,通過囊泡膜與軸突膜的融合,並在融合處出現裂口,使囊泡中的ACh全部進入接頭間隙。已故馮德培院士在30年代對神經-肌接頭的化學性質傳遞進行過重要的研究。
機能研究
對血糖機能
骨髓肌是具有收縮能力的肌細胞(由於其形狀成幼長的纖維狀,所以亦稱作肌纖維)所組成。任何的身體活動和體育活動,都是骨骼肌收縮的完成,直接影響人體的力量和耐力。諾貝爾生理學獎獲得者、義大利科學家DanielBovet經大量研究證實:骨骼肌在血糖利用方面作用極其重要,人體85%的血糖轉化和71%的糖元儲存由骨骼肌完成。骨髓肌——是具有收縮能力的組織之一,人體所有的活動幾乎都是由骨骼肌收縮來完成,其強弱直接影響人體的力量和耐力。人體85%以上的糖分是供給骨骼肌轉化成能量和體力的,是人體力量的主要能源。
專家研究發現,99.8%的糖尿病人骨骼肌出現弱化甚至萎縮現象,骨骼肌的弱化,一方面不能將糖分轉化為能量和體力,從而造成糖尿病人長期感覺疲憊、虛弱、乏力;另一方面由於糖分不能被骨骼肌完全利用,而在體內堆積,造成血糖升高。
同時,骨骼肌還是人體糖分主要的儲存場所,承擔了71%以上糖分的儲存,對人體血糖平衡具有極其重要的緩衝作用。一方面可以在血糖增多時將多餘糖分轉運存儲在骨骼肌中,避免糖分堆積在血液中使血糖升高;另一方面,當血糖過低時,骨骼肌釋放存儲的糖分,維持人體正常能量的需要,防止血糖過低。所以只有修補骨骼肌,才能打通人體用糖渠道,使血糖通過利用達到平衡,防止血糖淤積,平衡血糖代謝,防止併發症。
2006年,英國皇家糖尿病協會J.R.Kantor教授的糖尿病研究證實:自然界中有一種神奇物質——L阿拉伯糖,具有修補骨骼肌的顯著作用。2007年國際糖尿病聯盟(IDF)研究證實:L阿拉伯糖可以修補骨骼肌,有助於骨骼肌的恢復,加強骨骼肌對血糖的利用和存儲。國內由山東糖科院糖尿病研究中心的專家帶頭,經過科研和實驗證明了以上結論,即99.8%的糖尿病人骨骼肌出現弱化甚至萎縮現象,而且骨骼肌確實承擔了71%以上糖分的儲存,對人體血糖平衡具有極其重要的緩衝作用。
