
種屬概述
 蘭花蕉
蘭花蕉拉丁名:0rchidanthachinensisT.L.Wu
中文名:蘭花蕉
拉丁科名:Musaceae
中文科名:芭蕉科
保護級別:3
分布:廣東(信宜)、廣西(上思)
現狀:瀕危種。蘭花蕉自1932年首次在廣東信宜採得標本後,以後一直未在該地採到。目前僅知廣西上思十萬大山有分布。隨著森林過度砍伐,生態環境惡化,有漸趨絕滅的可能。
海拔下限(米):370
海拔上限(米):370
形態特徵
 蘭花蕉
蘭花蕉多年生草本,高約45厘米;根莖橫生。葉2列,葉片橢圓狀披針形,長22-30厘米,寬7-9厘米,頂端漸尖,基部楔形,稍下延,橫脈方格狀,稠密,乾時十分清楚;葉柄長14-18厘米。花自根莖生出,單生,苞片長圓形,長3.5-7厘米,位於花葶上部的較大,下部的較小。花大,紫色,萼片長圓狀披針形,長9.5 厘米,寬1.5-2厘米;唇瓣線形,長9厘米,基部寬8毫米,先端漸尖,具小尖頭,中部稍收縮;側生的2枚花瓣長圓形,長2厘米,先端有長5毫米的長芒;雄蕊5枚,花葯長1 厘米;子房頂端延長呈柄狀的部分長2厘米,花柱和花葯等長,柱頭3枚,其中1枚較長,長8 毫米,餘2枚稍短,先端具細鋸齒,背面具“V”形附屬物。果未見。花期:3月。
分布情況
產我國廣東、廣西,生於山谷中。模式標本采自廣東信宜。
品種分類
與“蘭花蕉”相關的種有:
海南蘭花蕉Orchidantha insularisT. L. Wu
保護級別
國家三級保護植物。
本種為芭蕉科蘭花蕉亞科在中國之代表種,數量少,分布極為局限,是稀有植物。對研究中國植物區系及芭蕉科分類系統均有研究價值。根莖民間用作清熱藥。
廣西上思十萬大山已建水源林區,應保護好那盪鄉紅旗林場後山蘭花蕉生長地自然林,嚴禁採挖。有關單位應大力繁殖、引種栽培。
種群研究
分布於廣東西南地區的蘭花蕉和廣西十萬大山的長萼蘭花蕉種群的樣地調查,分析蘭花蕉分布規律、種群結構、瀕危現狀及其瀕危原因,並提出相應的保護措施。蘭花蕉空間分布格局呈聚集型,種群規模小,致瀕的原因是環境破壞和自身繁殖能力下降。從形態學特徵上分析蘭花蕉及長萼蘭花蕉的5個葉部特徵,結果表明不管在種水平還是在居群水平,蘭花蕉及長萼蘭花蕉5個形態特徵差異都達顯著(P<0.05),其變異相對不太穩定,已出現一定的分化,且呈連續性。
生殖特性
蘭花蕉傳粉至受精的隔期大約為20天左右。開花時花葯已成熟散花粉,花粉主要是單核花粉,少數為2細胞花粉此時胚珠只是大孢子母細胞時期,開花後20-30天胚囊發育成熟。蘭花蕉花期為1個月,每朵花的花期為5天。蘭花蕉開花時柱頭已發育成熟,具有單細胞K毛狀腺組織,腺組織有許多粘液,在5天單花期中能夠接受花粉,萌發的花粉管經過花柱進入子房的延長部,花粉中的單核細胞和細胞隨之進入延長部。花凋謝後,單核細胞和2-細胞在延長部20天左右,繼續發育產生兩個精子與成熟胚囊進行雙受精。蘭花蕉子房延長部除了對子房具有保護作用外,還成為花粉繼續發育的場所,這是首次發現的功能。
形態解剖
蘭花蕉(0rchidanthachinensis)的子房室頂部閉合後向上延長成延長部,實心,但有花柱溝和隔膜蜜腺管通過,隔膜蜜腺管,可分為中央蜜腺管和三條側蜜腺管;中央蜜腺管位於三個心皮連線處,自子房室區下部產生,向上於延長部的部頂端終止;三條側管分別位於兩個心皮連線處,於子房室區近中部產生,開口於花柱基部。蘭花蕉子房室區與延長部均具6枚雄蕊的維管束系統,即3枚心皮背束的伴束與3枚隔膜束,近軸面1枚事膜向上進入唇瓣的維管束系統,位於唇瓣的中央,致使蘭花蕉僅具5枚功能雄蕊,唇瓣具雙重結構,本文還討論了蘭花蕉科的系統發育位置。
分布區年平均溫22,1月平均溫14,年平均降水量約1600毫米。本植物喜溫、濕,耐陰。多生於枯落葉層較厚,土壤深厚、肥沃、排水良好的溝谷山坡。花期3月,果期7月。
遺傳研究
 蘭花蕉
蘭花蕉採用簡單重複序列區間(ISSR,Inter-SimpleSequenceRepeat)分子標記技術,對采自廣東省的瀕危植物蘭花蕉(0rchidanthachinensisT.L.Wu)的7個居群137個個體進行遺傳變異分析。用10個ISSR引物共擴增出清晰譜帶101條,其中58條具有多態性,總多態位點百分率為57.43%.居群水平相對較低,多態位點百分率在6.93%-35.64%之間,平均為18.24%.經P0PGENE1.31數據處理,結果表明:在物種水平上Nei基因多樣性為0.1254±0.1686:Shann0n信息指數為0.2000±0.2429:Nei基因分化係數為0.5481,表明54.81%的遺傳變異分布在居群間,45.19%的遺傳變異分布在居群內。物種居群間的遺傳一致度在0.8855-0.9511之間。紅花潭是其最適合生境,建議在此建立自然保護區:鑒於蘭花蕉居群間出現了一定程度的分化,為最大限度地保護蘭花蕉的遺傳多樣性,建議在自然居群間進行相互移栽,以提高群體間的基因交流。
胚胎研究
 蘭花蕉染色體分析
蘭花蕉染色體分析蘭花蕉胚珠倒生,厚珠心,雙珠被,內外珠被共同構成珠孔周緣細胞組織僅一層。造孢細胞接發育成大孢子母細胞具直線形4分體和少數3分體。胚囊發育屬蓼型。成熟胚囊合點端變窄。形成肓囊。反足細胞存活時間較長胚珠具有珠心冠原和承珠盤結構。胚乳發育屬核型。種子脫落時,胚尚未分化:以上是蘭花蕉胚胎的主要特徵。
蘭花蕉(0rchidanthachinensisT.L.Wu)的胚珠倒生,具厚珠心和雙珠被。內外珠被形成珠孔。假種皮從外珠被的頂端發生。造孢時期,胚珠具有一層周緣細胞。造孢細胞發育成大孢子母細胞,大孢子母細胞減數分裂形成大孢子的線形四分體,少數三分體。合點大孢子具功能。胚囊發育屬蓼型。成熟胚囊的合點端狹長。胚珠具有珠心冠原和承珠盤。反足細胞壽命長。胚珠維管束屬於合點後多維管束類型。胚乳發育屬核型。種子脫落時,胚尚未分化出胚芽和胚根。所研究過的蘭花蕉屬植物中,只有兩種染色體倍性,即二倍體和六倍體。
就多倍體的起源而言,在其中間一般應有四倍體類群相聯繫,但到目前為止尚沒有觀察到四倍體種。遺憾的是在已發表的蘭花蕉屬植物中尚有部分種類無染色體數目的報導,如果這些種里不存在四倍體,那么以下的推測應是合理的:蘭花蕉屬植物尚還未發現的四倍體類群;但從蘭花蕉的生存環境來看,也不排除已經絕滅的可能。當然,因尚缺乏蘭花蕉居群的細胞學檢查,也不能排除該六倍體種為種內同源多倍體起源的可能。因此,該六倍體種的起源還需要進一步的研究。
花蕾長5cm左右時,子房胎座表皮下的細胞局部平周分裂,產生指狀突起,其前端發育成珠心,基部發育成珠柄。以後珠心與珠柄之間的細胞一側發育快,另一側發育慢,產生彎曲,從而形成倒生胚珠。在孢原細胞分裂時期,珠心基部發生一環狀的突起,後發育為內珠被接著在緊靠l珠被的基部外側又產生一個環狀突起發育成外珠被。外珠被較內珠被長,內外珠被形成珠孔珠被與珠心合併處為台點。胎座的維管束延伸到珠俑。
【胚珠結構】
珠心頂端的表皮下有1個體積火,細胞質濃,細胞核大的為孢原細胞,孢原細胞進行平周分裂,產生一個造孢細胞和一個周緣細胞。周緣細胞垂周分裂形成一層周緣組織細胞。珠孔端的周緣細胞不進行平周分裂,在二核胚囊時期逐漸消失。造孢細胞兩側的珠心細胞和周緣細胞,以及後部的珠心細胞進行有絲分裂增加珠心細胞,大孢子母細胞被包藏在發達的珠心組織中。蘭花蕉胚珠屬厚珠心型隨著胚囊的發育,胚囊周圍的珠心細胞從二核胚囊開始逐漸被消耗。受精後,胚和內胚乳的發育繼續消耗珠心細胞,最後僅剩少部分發育成外胚乳。珠心冠原和承珠盤珠孔端珠心表皮細胞從大孢子母細胞時期開始逐漸徑向伸長。細胞壁漸漸加厚胚囊成熟時,長形的表皮細胞排列成痢形的珠心冠原,這種結構有利花粉管進入胚囊;此外,盲囊周圍的細胞呈長形,體積大,細胞質少,壁加厚,此結構為承珠盤。
【珠被與假種皮】
胚珠成熟時,內珠被基部4層細胞,頂端9層細胞,呈棒狀外珠被13-16層細胞。外珠被的內表皮細胞呈正方形,細胞核太,圓形,染色深,以後發育成種子的一層內種皮。大孢子母細胞減數分裂時期,外珠被頂端局部內側細胞分裂較外側快,使外珠被頂端局部向外彎生,繼而發育成3-4條粗毛狀裂片的假種皮。
【珠孔】
大孢子母細胞時期內珠被漸漸靠攏,減數分裂時期合攏,但仍留有L隙,形成內珠孔一同時外珠被逐漸超過內珠被,也向中靠攏形成外珠孔。合點大孢子母細胞時期,核染色深,不斷分裂增加新的細胞胚囊成熟時,這些等徑的薄壁細胞已有12-15層,形成弧狀長條形。種子成熟時,這些薄壁細胞發育成合點區厚壁細胞群”。
【維管束】
胎座的維管束通過珠柄進人胚珠的合點,在合點區分支,形成8-10條維管束。蘭花蕉胚珠維管束屬合點後多維管束。
【大孢子的發生】
孢原細胞平周分裂產生兩個細胞,其下面一個為造孢細胞,造孢細胞直接發育成大孢子母細胞。當花蕾長到11cm長時,花開放,花葯成熟散花粉,此時的胚珠即是大孢子母細時期走孢子母細胞呈長圓形,細胞核大,圓形,細胞質濃。大孢子母細胞繼續發育增大。開花7d後,開始減數分裂,第一次分裂產生2個核,中間形成橫的分隔壁,即為二分體。二分體以同樣的方式分裂,形成直線形的四分體,形成4個大孢子有時也會出現三分體,這是因為二分體時,其中有一個細胞不再分裂,從而形成三分體。後珠L端的3個大孢子逐漸退化消失,合點端的大孢子繼續發育增大,最後占據整個胚囊腔,成為功能大孢子。
【雌配子體的發育與結構】
開花後12d,功能大孢子發生連續3次核的分裂。第一次分裂產生二核,它們分別移向兩極,不形成壁。二棱再一次分裂產生四核,在兩極各有二核。四核又各分裂一次形成八核,珠孔端四核,合點端四核,均沒有細胞壁,形成八核胚囊。隨著核的分裂,胚囊隨之擴大。隨後珠孔端和合點端各有一核移向胚囊中央,這兩個極核融合,形成中央細胞。珠孔端三核之問產生細胞壁形成三細胞,即卵細胞和兩個助細胞,兩個助細胞緊靠著,與卵細胞排成品字形的卵器。受精後助細胞退化消失,但有個別的助細胞可宿存到游離核形成幼期卵細胞充滿細胞質,隨發育產生液泡。合點端是三個反足細胞,染色較深。反足細胞存活時閭較長,在胚乳的游離核形成時,它們仍然存在。開花後20-30d胚囊發育成熟為七細胞胚囊。成熟胚囊前端和中間寬,前端圓形,合點端變窄,形成盲囊,反足細胞處在盲囊裡面。胚囊發育屬蓼型I。
【胚乳發育與胚的結構】
成熟胚囊進行雙受精。花粉管經過珠L和珠心冠原進八胚囊一個精子和卵融合形成合子;一個精子與中央細胞次生核融合成為三倍體的初生胚乳核。初生胚乳核經多次分裂,產生大量的核,但不形成壁。這些核是游離狀態分布在細胞質中,後因液泡的擴大,被擠到周圍發育後期在游離核之問產生細胞壁,變為細胞的胚乳。蘭花蕉胚乳的發育屬核型胚乳蘭花蕉的胚呈圓柱體。種子脫落時.胚乳已充分發育,而胚尚未分化少數具有胚芽原基和胚根原基的分生組織。蘭花蕉種子需要經過後熟,胚才能分化出胚芽和胚根。
【假種皮的發生】
蘭花蕉大孢子母細胞減數分裂時期,外珠被頂端局部內側細胞比外側細胞分裂快使外珠被頂端局部向外彎生。彎生的部分發育成假種皮這與Sachar等報導的姜科假種皮是由外珠做形成的看法一致。
染色體
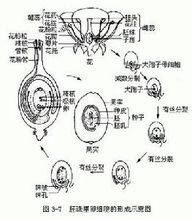 蘭花蕉胚珠結構
蘭花蕉胚珠結構首次報導了國產蘭花蕉屬植物2種1變種的體細胞染色體計數,其中蘭花蕉及其變種長萼蘭花蕉為2n=54,為六倍體:海南蘭花蕉為2n=18,為二倍體。分析了世界範圍內蘭花蕉屬植物染色體數及其倍性的地理學關係。
蘭花蕉體細胞染色體進行計數以來,至今僅3種蘭花蕉屬植物有細胞學資料,其體細胞染色體計數均為2n=18,到目前為止尚沒有觀察到有其它數目的報導。觀察了蘭花蕉屬植物兩種一變種的體細胞染色體數目,結果發現海南蘭花蕉為2n=18,但蘭花蕉及其變種長萼蘭花蕉則均為2n=54。馬來蘭花蕉是一個以x=9為基數的二倍體種,後來他(1993)研究的大花蘭花蕉都被確認為是以x=9為基數的二倍體種。我們研究的海南蘭花蕉2n=18,因此也應該是二倍體種,而蘭花蕉和長萼蘭花蕉則應是x=9的六倍體。有關結果列表比較。從研究結果看,長萼蘭花蕉的染色體數目與蘭花蕉的一樣,都為2n=54,因此染色體資料支持吳德鄰的觀點,即將長萼蘭花蕉作為蘭花蕉的變種處理。
栽培方法及注意事項
 蘭花蕉
蘭花蕉春夏間切割帶芽的橫生根莖,進行無性繁殖,植於有適當蔭蔽、土壤肥沃、排水良好的溝谷山坡。
繁殖
用分株繁殖。當秋末蕉葉凋萎,剪去枯葉,壅士護根。殘留莖桿用稻草從莖桿基部向上包紮。到次年4月上旬再將稻草解除,當根上長出許多幼株時,可行分株繁殖。移栽時於坑中施入有機肥為底肥。
栽培
生長期間應隨時剪去黃葉,以燒徒耗養分,並影響美觀。同一地點,栽植過久,易產生愛滿不復現象,應更換栽植地點。平時不用特別仔細管理。
蘭花蕉最適宜植於小型庭院的一角或窗前牆邊,假山之畔。不宜成行栽植,宜散點或幾株叢植,綠蔭如蓋,炎夏中令人頓生清涼之感。蘭花蕉是多年生草本植物,體積龐大,適於庭院栽培。倘若設法矮化,便可移為居室盆栽。六月間將母株邊上新發的小株分割下來,選基本成單株狀,最好有淺根系的,將切口用草木灰塗勻以防腐爛。芭蕉擇土不嚴,但需排水性好,可選沙子20%、土60%、木屑20%混拌。用盆以直徑為60?80厘米左右的大盆或缸為宜,深度應不低於70厘米。定植後要少澆水,噴灑表土即可。待有新葉長出,可逐步增加噴水量及見光量。養護期間要注意保持濕潤,在高溫高濕的環境下才能生長良好。同時也要適當延長光照時間,以防枝葉徒長。
主要價值
根狀莖:用於斑疹不退,煩熱,咽喉腫痛。
瀕危種。本種為蕉科蘭花亞科在中國之代表種,數量少,分布極為局限,是稀有植物。對研究我國植物區系及蕉科分類系統均有研究價值。根莖民間用作清熱藥。
中國珍稀瀕危植物3
| 在自然界中,凡是有生命的機體,均屬於生物。生物應分為幾個界,把行固著生活和自養的生物稱為植物界,簡稱植物。植物的特點是具有光合作用的能力——就是說它可以藉助光能及動物體內所不具備的葉綠素,利用水、礦物質和二氧化碳生產食物。釋放氧氣後,剩下葡萄糖——含有豐富能量的物質,作為植物細胞的組成部分。植物通常是不運動的,因為它們不需要尋找食物。我們一起來認識這些植物吧! |
