主要特徵
常綠或單葉喬木,主幹發達,具頂枝起源葉,葉呈針形、線形、披針形、刺形或鱗片形,
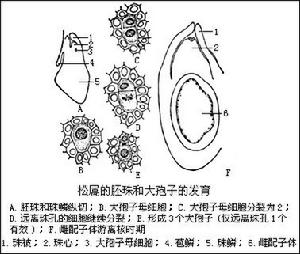 胚珠及大孢子發育
胚珠及大孢子發育松柏綱植物精細胞無纖毛,這種性狀在裸子植物系統發育上較蘇鐵綱和銀杏綱進化.
繁殖方式
松柏綱植物的生殖器官分為雄球花和雌球花兩種。雄球花會產生花粉;雌球花在其木質鱗片之
 松柏盆景
松柏盆景生活史
以最普遍而研究最詳盡的松科(Pinaceae)的松屬(Pinus)為代表,介紹如下。
 松柏綱
松柏綱(一)孢子體
松屬的孢子體為高大多年生常綠喬木,單軸分枝,主幹直立,旁枝輪生,具長枝和短枝。網狀中柱,90—95%由管胞組成,樹脂道約占1%,木射線約占6%。長枝上生鱗葉,腋內生短枝,短枝極短,頂生1束針形葉,每束通常2、3、5個葉,基部常有薄膜狀的葉鞘8—12枚(由芽鱗變成)包圍,葉內有1或2條維管束和幾個樹脂道。
孢子葉球單性,同株。小孢子葉球排列如穗狀,生在每年新生的長枝條基部,由鱗片葉腋中生出。每個小孢子葉球有1個縱軸,縱軸上螺鏇狀排列著小孢子葉,小孢子葉的背面(遠軸面)有1對長形的小孢子囊。小孢子囊內的小孢子母細胞,經過兩次的連續分裂(其中一次為減數分裂),形成4個小孢子(花粉粒)。小孢子有2層壁,外壁向兩側突出成氣囊,能使小孢子在空氣中飄浮,便於風力傳播。
大孢子葉球1個或數個著生於每年新枝的近頂部,初生時呈紅色或紫色,以後變綠,成熟時為褐色。大孢子葉球是由大孢子葉構成的,大孢子葉也是螺鏇狀排列在縱軸上的,但它們不是簡單的孢子葉,而是由兩部分組成:下面較小的薄片稱為苞鱗;上面較大而頂部肥厚的部分稱為珠鱗,也叫果鱗或種鱗,一般認為珠鱗是大孢子葉,苞鱗是失去生殖能力的大孢子葉。在松科各屬植物苞鱗和珠鱗是完全分離的,每1珠鱗的基部近軸面著生2個胚珠,胚珠由1層珠被和珠心組成,珠被包圍著珠心,形成珠孔。珠心即大孢子囊,中間有1個細胞發育
 松柏綱
松柏綱成大孢子母細胞,經過兩次連續分裂(其中一次是減數分裂),形成4個大孢子,排列成1列稱為“鏈狀四分體”。通常只有合點端的1個大孢子發育成雌配子體,其餘3個退化。
(二)雄配子體
雄配子體是1個大為減退了的結構,只由少數幾個細胞構成。
小孢子(單核時期的花粉粒)是雄配子體的第一個細胞,小孢子在小孢子囊內萌發,細胞分裂為2,其中較小的1個是第一個原葉體細胞(營養細胞),另1個大的叫胚性細胞,胚性細胞再分裂為2,即第二原葉細胞及精子器原始細胞(中央細胞),精子器原始細胞再分裂為2,形成管細胞和生殖細胞。成熟的雄配子體有4個細胞:2個退化原葉體細胞、1個管細胞和1個生殖細胞。
(三)雌配子體
由大孢子發育而成。因此,大孢子是雌配子體的第一細胞,它在大孢子囊(珠心)內萌發,進行游離核分裂,形成具16—32個游離核,不形成細胞壁。雌配子體的四周具1薄層細胞質,中央為1個大液泡,游離核多少均勻分布於細胞質中,當冬季到來時,雌配子體即進入休眠期。翌年春天,雌配子體重新開始活躍起來,游
 松的生活史
松的生活史(四)傳粉與受精
傳粉在晚春進行,此時大孢子葉球軸稍為伸長,使幼嫩的苞鱗及珠鱗略為張開。同時,小孢子囊背面裂開一條直縫,處於雄配子體階段的花粉粒,借風力傳播,飄落在由珠孔溢出的傳粉滴中,並隨液體的乾涸而被吸入珠孔。這時大孢子葉球的珠鱗又繼續閉合。雄配子體中的生殖細胞分裂為2,形成1個柄細胞及1個體細胞,而管細胞則開始伸長,迅速長出花粉管。但這時大孢子尚未形成雌配子體,花粉管進入珠心相當距離後,即暫時停止伸長,直到第二年春季或夏季頸卵器分化形成後,花粉管才再繼續伸長,此時體細胞再分裂形成2個精子(不動精子)。受精作用通常是在傳粉以後13個月才進行,即傳粉在第一年的春季,受精在第二年夏季。這時大孢子葉球已長大並達到或將達到其最大體積,頸卵器已完全發育。當花粉管伸長至頸卵器,破壞頸細胞到達卵細胞處,其先端隨即破裂,2個精子、管細胞及柄細胞都一起流入卵細胞的細胞質中,其中1個具功能精子隨即向中央移動,並接近卵核,最後與卵核結合形成受精卵,這個過程稱受精。
(五)種子
松屬的胚胎髮育過程,頗複雜,具明顯的階段性,通常可分原胚階段、胚胎選擇階段、胚的器官和組織分化階段、胚的成熟階段。但這些階段是按順序連續發育的,是相互聯繫和相互制約的。
1.原胚階段:從受精卵分裂開始到細胞型原胚的形成,先後經過游離核的分裂、細胞壁的產生和原胚的形成。
受精卵接連進行3次游離核的分裂,形成8個游離核,這8個游離核在頸卵器基部排成上、下兩層,每層4個,細胞壁即在此時形成,但上層4個細胞的上部不形成胞壁,使這些細胞的細胞質與卵細胞質相通,稱為開放層;下層4個細胞稱為初生胚細胞層。接著開放層和初生胚細胞層各自再分裂1次,形成4層,分別
 銀杉
銀杉2.胚胎選擇階段:胚柄系統的發育和多胚現象的產生是這個階段的主要特徵。原胚的4層細胞從上到下,第一層(上層),初期有吸收作用,不久即解體;第二層蓮座層,分裂數次之後消失;第三層胚柄層,它的4個細胞稱為初生胚柄,不再分裂,但伸長;第四層胚細胞層的胚細胞,在胚柄細胞繼續延長的同時,緊接著後面的胚細胞進行分裂並伸長,稱為次生胚柄,由於胚柄和次生胚柄(胚管)迅速伸長,形成多回捲曲的胚柄系統。而胚細胞層的最前端的細胞發育成胚的本身,但它們不組成1個胚,而在縱面彼此分離,各個單獨發育成胚,稱為多胚現象。常見的多胚現象有兩種:一種是簡單多胚現象,即在同一個胚珠內有2個以上的頸卵器的卵細胞,可以同時受精,因而在胚胎髮育的早期,可以產生2個以上的原胚;另一種是裂生多胚現象,即由1個受精卵形成的4個胚細胞,分別單獨發育成為4個幼胚。
在胚胎髮育過程中,通過胚胎選擇,通常只有1個(很少2個或更多)幼胚正常分化、發育,成為種子中成熟的胚。
大多數松柏類植物具有2個以上的頸卵器,所以簡單多胚現象是普遍發生的,至於裂生多胚現象,則僅限於松柏類植物的幾個屬可見。
3.胚的器官和組織分化階段:胚在進一步的發育中成為1個伸長的圓柱體。這個圓柱體的近軸區(基部)同胚柄系統相接,主要是橫分裂,細胞略大,形成較規則的行列,進而發育成根端和根冠組織;而在遠軸區內,細胞分裂似無特定的方向,細胞較小,由這些細胞進一步分化,最後分裂出下胚軸、胚芽和子葉。
4.胚的成熟階段:成熟的胚包括胚根、胚軸(胚莖)、胚芽和子葉(通常7—10枚)。包圍此胚的雌配子體(胚乳)繼續生長,最後珠心僅遺留一薄層。珠被發育成種皮,種皮分為3層:外層肉質(不發達)、中層石質、內層紙質。
胚、胚乳、種皮構成種子。
裸子植物的種子是由3個世代的產物組成的,即胚是新的孢子體世代(2n);胚乳是雌配子體世代(n);種皮是老的孢子體(2n)。受精後,大孢子葉球繼續發育,珠鱗木質化而成為種鱗,種鱗頂端擴大露出的部分為鱗盾,鱗盾中部有隆起或凹陷的部分為鱗臍,珠鱗的部分表皮分離出來形成種子的附屬物即翅,以利風力傳播。種子萌發時,主根先經珠孔伸出種皮,並很快產生側根,初時子葉留在種子內,從胚乳中吸取養料,隨著胚軸和子葉的不斷發展,種皮破裂,子葉露出,而隨著莖頂端的生長,產生新的植物體。
下屬分類
松柏綱植物是現代裸子植物中數目最多、分布最廣的類群。現代松柏綱植物有44屬,約400餘種,
 金錢松
金錢松(一)松科(Pinaceae)
喬木,稀灌木,大多數常綠。葉條形或針形:條形葉扁平,稀呈四棱形,在長枝上螺鏇狀散生,在短枝上簇生;針形葉常2—5針成束,著生於極度退化的短枝頂端,基部包有葉鞘。孢子葉球單性同株,小孢子葉球,具多數螺鏇狀著生的小孢子葉,每個小孢子葉有2個小孢子囊,小孢子多數有氣囊;大孢子葉球,由多數螺鏇狀著生的珠鱗與苞鱗所組成,每珠鱗的腹面(上面)具兩枚倒生的胚珠,背面(下面)的苞鱗與珠鱗分離(僅基部結合),花後珠鱗增大發育成種鱗。球果直立或下垂。種子通常有翅;胚具2—16枚子葉。染色體:X=12、13、22。
本科是松柏綱植物中最大而且在經濟上又是重要的1科,有10屬,約230餘種,多產於北半球。中國有10屬,113種(包括引種栽培24種),分布遍於全國,絕大多數都是森林樹種和用材樹種,以及許多還是特有屬和孑遺植物。
本科植物以具針形葉或條形葉,葉及種鱗螺鏇狀排列,種鱗與苞鱗離生,每種鱗具2粒種子等為特色。
(二)杉科(Taxodiaceae)
喬木。葉螺鏇狀排列,同一樹上的葉同型或二型;孢子葉球單性同株,小孢子葉及珠鱗螺鏇狀排列(僅水衫的葉和小孢子葉、珠鱗對生),小孢子囊多於2個(常3—4個),小孢子無氣囊,珠鱗與苞鱗多為半合生(僅頂端分離),珠鱗的腹面基部有2—9枚直立或倒生胚珠。球果當年成熟,種鱗(或苞鱗)扁平或盾形,木質或革質,能育種鱗有2—9粒種子,種子周圍或兩側有窄翅。染色體:X=11、33。
 水杉
水杉本科有10屬,16種,主要分布於北半球。中國產5屬,7種,引入栽培4屬,7種,分布於長江流域及秦嶺以南各省區。
本科植物以種鱗和苞鱗半合生,種鱗具2—9粒種子,葉披針形、鑽形、條形或鱗狀,互生,螺鏇狀排列或2列(除水杉屬對生外),小孢子無氣囊等為特色。
(三)柏科(Cupressaceae)
常綠喬木或灌木。葉互動對生或輪生,稀螺鏇狀著生,鱗形或刺形,或同一樹上兼有兩型葉。孢子葉球單性,同株或異株。小孢子葉互動對生,小孢子囊常多於2個,小孢子無氣囊。珠鱗交又對生或3—4片輪生,珠鱗腹面基部有1至多枚直立胚珠,苞鱗與珠鱗完全合生。球果通常圓球形,種鱗盾形,木質或肉質,熟時張開或肉質合生呈漿果狀。種子兩側具窄翅或無翅,或上端有1長1短的翅。染色體:X=11。
本科共22屬,約150種,分布於南、北兩半球。中國產8屬,29種,分布幾遍全國,另引入栽培1屬,15種。
本科植物以葉對生或輪生,具兩型葉,種鱗和苞片完全合生,珠鱗互動對生或3—4片輪生,胚珠直立等為特色。
代表植物
銀杉
屬裸子植物,松科。國家一級保護植物。別名衫公子,是一種高十至二十幾米的常綠喬木。它是
 華北落葉松
華北落葉松常綠喬木,具開展的枝條,高達24米,胸徑通常達40厘米,稀達85厘米;樹幹通直,樹皮暗灰色,裂成不規則的薄片;小枝上端和側枝生長緩慢,淺黃褐色,無毛,或初被短毛,後變無毛,具微隆起的葉枕;芽無樹脂,芽鱗脫落。葉螺鏇狀排列,輻射狀散生,在小枝上端和側枝上排列較密,線形,微曲或直通常長4~6厘米,寬2.5~3毫米,先端圓或鈍尖,基部漸窄成不明顯的葉柄,上面中脈凹陷,深綠色;無毛或有短毛,下面沿中脈兩側有明顯的白色氣孔帶,邊緣微反卷,橫切面上有2個邊生樹脂道;幼葉邊緣具睫毛。雌雄同株,雄球花通常單生於2年生枝葉腋;雌球花單生於當年生枝葉腋。球果兩年成熟,卵圓形,長3~5厘米,直徑1.5~3厘米,熟時淡褐色或栗褐色;種鱗13~16枚,木質,蚌殼狀,近圓形,背面有短毛,腹面基部著生兩粒種子,宿存;苞鱗小,卵狀三角形,具長尖,不露出;種子倒卵圓形,長5~6毫米,暗橄欖綠色,具不規則的斑點,種翅長10~15毫米。
雲杉
常綠喬木。樹幹端直。中國是雲杉屬植物最多的國家,共16種9變種,分布於東北、華北、西北、西南地區及台灣的山地及亞高山地帶,尤以橫斷山地區種類最多,木材蓄積量豐富。雲杉屬植物最早的化石發現於美國西部及日本的晚白堊紀地層中。第三紀末至第四紀更新世因紅皮雲杉全球性氣溫下降的影響,使雲杉從高緯度和高海拔地區向低緯度和低海拔地區擴展,種類增加。後隨冰川的退縮和氣溫的回升,分布區又逐漸縮減,繁衍至今,形成了現代的分布格局。在中國秦嶺以南及東南部的平原和西南部的低山地區的晚更新世沉積物中發現了雲杉的花粉及球果。
許多雲杉樹的特徵具有下垂的枝,整顆樹層層排列。直挺的針狀葉呈螺鏇形圍繞著莖,長度為2~3厘米。葉從椿處長出來,如果脫落,椿便會保留下來。木質球果懸吊著,卵石形成圓柱形,有向內彎曲的
 雲杉
雲杉水杉
落葉喬木,高達35一41.5米,胸徑達1.6-2.4米;樹皮灰褐色或深灰色,裂成條片狀脫落;小枝對生或近對生,下垂。葉互動對生,在綠色脫落的側生小枝上排成羽狀二列,線形,柔軟,幾無柄,通常長1.3一2厘米,寬1.5一2毫米,上面中脈凹下,下面沿中脈兩側有4-8條氣孔線。雌雄同株,雄球花單生葉腋或苞腋,卵圓形,互動對生排成總狀或圓錐花序狀,雄蕊互動對生,約20枚,花葯3,花絲短,藥隔顯著;雌球花單生側枝頂端,由22-28枚互動對生的苞鱗和珠鱗所組成,各有5一9胚珠。球果下垂,當年成熟,果實藍色,可食用,近球形或長圓狀球形,微具四棱,長1.8-2.5厘米;種鱗極薄,透明;苞鱗木質,盾形,背面橫菱形,有一橫槽,熟時深褐色;種子倒卵形,扁平,周圍有窄翅,先端有凹缺。
