
概念
細胞工程(Cellengineering):
是指套用現代細胞生物學、發育生物學、遺傳學和分子生物學的理論與方法,按照人們的需要和設計,在細胞水平上的遺傳操作,重組細胞結構和內含物,以改變生物的結構和功能,即通過細胞融合、核質移植、染色體或基因移植以及組織和細胞培養等方法,快速繁殖和培養出人們所需要的新物種的生物工程技術。
細胞工程與基因工程一起代表著生物技術最新的發展前沿,伴隨著試管植物、試管動物、轉基因生物反應器等相繼問世,細胞工程在生命科學、農業、醫藥、食品、環境保護等領域發揮著越來越重要的作用。
21世紀合成生物學的發展,採用計算機輔助設計、DNA或基因合成技術,人工設計細胞的信號傳導與基因表達調控網路,乃至整個基因組與細胞的人工設計與合成,從而刷新了基因工程與細胞工程技術,並將帶來生物計算機、細胞製藥廠、生物煉製石油等技術與產業革命。
特點
1.前沿性:現代生物技術的熱點
人類染色體圖片
人類染色體圖片
2.爭議性:新技術給倫理道德帶來的衝擊
3.綜合性:多學科交叉
4.套用性:工程類課程,重在產品與技術
研究內容
動植物細胞與組織培養
細胞融合(新的物種或品系、單克隆抗體)
細胞核移植(無性繁殖、克隆動物)
染色體工程(多倍體育種,例:八倍體小黑麥)
胚胎工程(優良品種、試管嬰兒)
幹細胞與組織工程(胚胎幹細胞、組織幹細胞)
轉基因生物與生物反應器(轉基因動物、轉基因植物)
核心技術
核心技術:細胞培養與繁殖
目的:獲得新性狀、新個體、新物質或產品
發展簡史
 染色體圖片
染色體圖片細胞的發現
1665年,英國人胡克(Hooke)利用自己設計的顯微鏡第一次觀察到了細胞。
細胞理論的提出
1838年,施萊登(Schleiden)發表“植物發生論”,認為無論怎樣複雜的植物都由細胞構成。
1839年,施旺(Schwann)發表“關於動植物結構和生長一致性的顯微研究”。提出“細胞學說”(CellTheory)。之後,德國科學家魏爾肖(Virchow)補充了細胞學說,認為所有的細胞都來自於已有的細胞分裂。細胞學說的建立揭示了生物界的統一性和生命的共同起源,是19世紀自然科學的三大發現之一。
細胞與組織培養
1902,Haberlandt,植物細胞全能學說。
1907,Harrison,蛙胚神經細胞突起,無菌操作技術。
1912,Carrel,雞胚心肌組織塊長期傳代培養。
1940,Earle,首創單個細胞克隆培養,建立小鼠結締組織
L細胞系,並在1951年開發了人工培養液。
細胞融合
1975,CesarMilstein與GeogerKohler合作,羊紅細胞免疫過的小鼠脾細胞與小鼠骨髓瘤細胞融合,得到既能體外無限繁殖又能產生特異性抗體的雜交瘤細胞。
胚胎工程
1978年,英國劍橋大學生理學家羅伯特·愛德華採用胚胎工程技術成功培育出世界首例試管嬰兒-路易絲-布朗。
轉基因生物
1983年Palmiter和Brinster將大鼠生長激素基因轉入小鼠,生產出生長速度極快的碩鼠。
1987年Gordon獲得分泌組織纖溶酶原激活因子tPA的轉基因小鼠。
細胞核移植
1938年,德國胚胎學家Spemann提出胚胎細胞核移植到去核卵母細胞中可發育為新胚胎。
1997年,“多莉”羊的誕生標誌著哺乳動物的體細胞核克隆時代到來。
種類
染色體工程
將一種生物的特定染色體,按照人們的意圖予以消除、添加、或同別的生物的染色體置換、或改造的技術。目前基因工程的操作技術多限於單個或少數基因在大腸桿菌等微生物中的表達。為了改變真核細胞的遺傳性和控制高等生物的生命活動,還必須研究和開發染色體工程,建立一種新的技術體系,把所需的基因或染色體片段整合到染色體的任意位置,並能將有關遺傳信息在細胞分裂中一代又一代的傳遞下去。目前這方面的工作還處於起步階段。
動物細胞的染色體工程
又稱為染色體轉導,或染色體介導的基因的轉移。染色體轉導術,目前有兩類,其一,稱為微細胞轉移術。套用低濃度秋水仙素長時間處理可使細胞微核化,經去核處理後,可得到只含相當於幾個乃至一個染色體的微細胞。微細胞被導入完整細胞以後仍顯示RNA合成,因而微核編碼的基因信息可望在微細胞異核體內表達出來。如小鼠的微細胞可被導入至另一品系的小鼠或倉鼠乃至人的HeLa細胞內。電泳檢測顯示存在著小鼠基因型的大分子物質,如脂酶D、嘌呤核苷磷酸化酶和肽酶 B。已知前兩種酶的結構基因定位於小鼠的第14號染色體上。提示小鼠的該號染色體已進入宿主細胞內並行使其功能。 另一種方法是先誘發細胞同步分裂,繼用秋水仙素阻抑細胞分裂於中期,再破碎細胞,通過離心收集大量的中期染色體。有人把此法得到的人或倉鼠的中期染色體轉移到小鼠細胞內,並探查到有特異的供體基因的功能產物, 動物的染色體工程與育種
動物的染色體工程與育種 植物的染色體工程與育種
植物的染色體工程與育種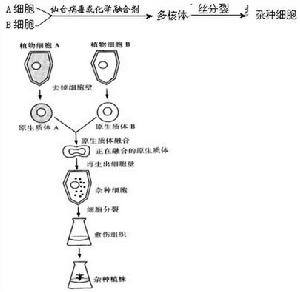 生物熱點 細胞工程
生物熱點 細胞工程染色體組工程的方法多倍體的誘發 自1937年發現了用秋水仙素誘發多倍體的方法以來,一般常用藥劑(秋水仙素、富民隆等),也可用高溫處理來誘發多倍體。其法是把植物的種子或幼芽浸在 0.05~0.2%的秋水仙素水溶液中,處理24~96小時即可得到很好的效果。例如四倍體西瓜、甜菜、玉米和百合等都是用此法獲得的(圖2)。 現在,由於原生質體分離技術的發展,也可從原生質體的融合得到多倍體。例如用聚乙二醇作誘導融合劑處理胡蘿蔔原生質體後,得到了頻率相當高的四倍體和六倍體植株。這是來源於二個或三個原生質體融合的結果。 單倍體的誘發60年代以來,子房、花葯或花粉離體培養成功,很易從大孢子、卵細胞或小孢子等得到單倍體植株。其法是將一定時期的花葯或子房移植到特定的培養基上培養。待生長愈傷組織或胚狀體後,再移到分化培養基上,分化出苗和根,長成完整的小植株即可移到盛有土壤的盆中繼續栽培到開花。單倍體植物一般不能結實或僅結少量種子。 此外,還可用遠緣雜交,X射線或紫外線照射,化學藥品如馬來醯肼、甲苯胺藍、氯黴素等以及異源胞質等方法都能誘導單倍體產生。1970年有人又用大麥與球莖大麥雜交後染色體消除的方法,產生高頻率的單倍體,有的可高達68.5%。在雜交後,球莖大麥的7個染色體就消除在胚中,留下的是大麥的7個染色體,成為單倍體的胚及小植株。經秋水仙素處理後,染色體加倍形成純合二倍體。 染色體組工程的套用誘導多倍體在植物育種上的套用是有限度的。由於作物類型不同,對多倍性誘變反應也不同。原來的倍性水平、染色體組的結構、繁殖方式、多年生性、植株實用部位,所有這些都關係到育種的成敗。最適宜用染色體加倍方法改良的作物應該具有:①染色體數目較少,②以收穫營養體為主,③異花授粉,④多年生和營養繁殖的習性等條件。這些都是多倍體育種獲得成功的先決條件。 細胞質工程研究真核細胞的核、質相互關係以及細胞器,胞質基因的轉移等細胞拆合的技術,所以又叫細胞拆合工程。主要研究內容是細胞質的置換。過去在植物上置換的方法是進行連續回交。例如,為了研究柳葉菜屬的細胞質遺傳,曾連續回交了二十五代,結果還不能把全部母核替代出來。現在由於核移植和原生質體的分離方法的改進,推進了這項工程的進展。 細胞質工程的方法去核和核移植 動物細胞核的移植一般都用顯微操作器進行。50年代初期,美國生物學家R.布里格斯和T.金首先成功地把豹蛙囊胚期細胞的細胞核移植到去核的蛙卵,並能正常發育。後來,英國J.B.格登把爪蟾蝌蚪腸上皮細胞核移植到去核卵內,能發育到有生殖能力的成體。中國童第周等還成功地進行金魚類異種、異屬之間的核移植實驗(見細胞分化)。70年代以來,體外培養的動物細胞的去核,是先用細胞鬆弛素B處理細胞,再高速離心使細胞核與細胞質分開。分離出來的核,帶有少量胞質並圍有質膜,稱為“核體”或“小型細胞”。核體能重新再生其胞質部分,繼續生長、分裂。去核後的胞質部分,仍由膜所包圍,即為“胞質體”或“去核細胞”(圖3)。秋水仙素及其衍生物和長春新鹼等也能誘發某些哺乳類細胞排核。目前製備胞質體和核體的方法目臻完善,純度可達99%左右。胞質體約可存活18~36小時。 植物細胞核的移植,在低等植物如單細胞傘藻,可把新鮮材料的假根切下,放在玻片上用玻棒擠壓,使細胞的內含物壓出在一滴適合的培養液中,反覆沖洗幾次,然後在顯微鏡下觀察,一直到核周圍無細胞質為止。離心分離後待用。高等植物如矮牽牛、天仙子、菸草、番茄等原生質體核的分離,可先在懸浮的原生質體中用蒸餾水將懸液沖淡一半,約30分鐘後,原生質體破裂,放出細胞核與葉綠體,就可在0.6M蔗糖液中離心和收集核,然後存放在一定的培養液中待用。1978年以來又借用動物細胞去核的藥劑細胞鬆弛素B來處理原生質體,加上高速離心,使原生質體分離成二部分,即:無核原生質體和小原生質體。開闢了去植物細胞核甚至去部分染色體的新途徑。
 細胞工程
細胞工程 動物細胞融合過程
動物細胞融合過程植物方面 用等滲密度梯度高速離心後,也可得到兩種亞原生質體,①在低密度範圍內可得到胞質體(去核原生質體);②在高密度中,可得到小原生質體(核質體)。現在已能自玉米、菸草和胡蘿蔔細胞得到這兩種亞原生質體。生化實驗證明:去核原生質體代謝作用很低,而小原生質體由於減少了表面積和體積(僅及原生質體的10~15%),因此,攝取物質快,合成蛋白質也快,培養時發育迅速,是一種研究核質關係的好材料。由於這項工作才開始,迄今尚無明顯結果。 細胞融合工程細胞融合是指用自然或人工的方法,使兩個或幾個不同的細胞融合成一個細胞的過程。細胞融合的結果,一個細胞中含有兩個不同的細胞核,則稱為異核體;隨後的有絲分裂中,來自不同細胞核的染色體可能合併到一個結合核內。因此,又稱為體細胞雜交。細胞融合的範圍很廣,從種內、種間、屬間、科間一直到動、植物兩界之間都進行了嘗試。在植物方面,由於各類細胞具有全能性,在菸草、矮牽牛、胡蘿蔔等種間雜種,馬鈴薯和番茄、曼陀羅和顛茄、菸草和矮牽牛等屬間雜種都已獲得了再生植株。在動物方面人和鼠體細胞雜交,雖然不能長成一個新個體,但能作基因定位的材料。因此,這項新技術,在理論研究和工、農、醫方面的套用,均有廣闊的前景。細胞融合技術的發展,歷史很短。自1960年在體外培養中發現雜種細胞以來,僅20多年。1965年岡田善雄等和H.哈里斯等各自用滅活的仙台病毒誘導產生了第一個種間異核體。1970年已套用人與鼠的細胞雜交系統地進行了人類染色體基因的定位工作。在植物方面,1960年E.C.科金首先使用纖維素酶分離番茄幼根的原生質體獲得成功。1970年他們又成功地使種間原生質體融合在一起。1972年P.S.卡爾森等又從融合的原生質體獲得了第一株種間細胞雜種。到1980年為止,種間融合的再生植株已有16種之多。 細胞融合的方法 動物細胞雜交或細胞融合 將兩個不同種的親本細胞A和B,以滅活的仙台病毒或聚乙二醇(PEG)為融合誘導劑,使A和B兩細胞融合成為一個具兩個遺傳性不同核的異核體(如遺傳性相同的核融合在一起叫同核體)。隨後異核體經有絲分裂成為兩個具有A和B兩親本的雜種融合核。AB雜種經多次分裂,B親本的染色體會逐漸減少到一個或完全消失(圖6)。 植物體細胞雜交 ①原生質體的分離。植物細胞之間有果膠質粘連,每個細胞之外還有一層纖維素組成的壁,因此,在分離原生質體時,首先要在一定濃度的酶液(果膠酶與纖維素酶)中保溫,消去果膠質與纖維素後才能使原生質體分離出來。②原生質體的融合。不同種之間原生質體的融合,須選用一種融合誘導劑(聚乙二醇,或高鈣CaCl2.2啹O,0.05M溶於甘露醇 0.4M和pH10.5)誘導融合。它們的誘導率可達20~50%。③雜種細胞的選擇與培養。細胞融合後要把雜種細胞選擇出來。一般都利用各種生化指標和遺傳標記來選擇和鑑定。例如,使用天然的或人工誘變的突變體,如白化苗、營養缺陷型、抗藥性突變體等,或根據不同材料對激素敏感性不同,生長差異等,來設計適合的選擇系統。如果融合的原生質體一個是白化,另一個具葉綠體,就可用機械的方法,把融合的細胞在倒置顯微鏡下把它們挑選出來進行培養。這些細胞培養到各個發育階段,如愈傷組織、分化苗和根,都需要更換培養基,才能使它們順利地再生成植株。
套用
細胞工程作為科學研究的一種手段,已經滲入到生物工程的各個方面,成為必不可少的配套技術。在農林、園藝和醫學等領域中,細胞工程正在為人類做出巨大的貢獻。
1.糧食與蔬菜生產利用細胞工程技術進行作物育種,是迄今人類受益最多的一個方面。我國在這一領域已達到世界先進水平,以花葯單倍體育種途徑,培育出的水稻品種或品系有近百個,小麥有30個左右。其中河南省農科院培育的小麥新品種,具有抗倒伏、抗鏽病、抗白粉病等優良性狀。
 土豆和番茄的融合
土豆和番茄的融合在果樹、林木生產實踐中套用細胞工程技術主要是微繁殖和去病毒技術。幾乎所有的果樹都患有病毒病,而且多是通過營養體繁殖代代相傳的。用去病毒試管苗技術,可以有效地防止病毒病的侵害,恢複種性並加速繁殖速度。目前,香蕉、柑橘、山楂、葡萄、桃、梨、荔枝、龍眼、核桃等十餘種果樹的試管苗去病毒技術,已基本成熟。香蕉去病毒試管苗的微繁殖技術已成為產業化商品化的先例之一。因為香蕉是三倍體植物,必須通過無性繁殖延續後代,傳統方法一般採用芽繁殖,感病嚴重,繁殖率低;而採用去病毒的微繁殖技術不僅改進了品質,畝產量約提高30%~50%,很容易被蕉農接受。
 細胞融合抗鹽鹼耐乾旱葡萄
細胞融合抗鹽鹼耐乾旱葡萄自1975年英國劍橋大學的科學家利用動物細胞融合技術首次獲得單克隆抗體以來,許多人類無能為力的病毒性疾病遇到了剋星。用單克隆抗體可以檢測出多種病毒中非常細微的株間差異,鑑定細菌的種型和亞種。這些都是傳統血清法或動物免疫法所做不到的,而且診斷異常準確,誤診率大大降低。例如,抗B型肝炎病毒表面抗原(HBsAg)的單克隆抗體,其靈敏度比當前最佳的抗血清還要高100倍,能檢測出抗血清的60%的假陰性。
 B16黑色素瘤與活化B淋巴細胞融合瘤細胞免疫原性的改變
B16黑色素瘤與活化B淋巴細胞融合瘤細胞免疫原性的改變目前,人工受精、胚胎移植等技術已廣泛套用於畜牧業生產。精液和胚胎的液氮超低溫(-196攝氏度)保存技術的綜合使用,使優良公畜、禽的交配數與交配範圍大為擴展,並且突破了動物交配的季節限制。另外,可以從優良母畜或公畜中分離出卵細胞與精子,在體外受精,然後再將人工控制的新型受精卵種植到種質較差的母畜子宮內,繁殖優良新個體。綜合利用各項技術,如胚胎分割技術、核移植細胞融合技術、顯微操作技術等,在細胞水平改造卵細胞,有可能創造出高產奶牛、瘦肉型豬等新品種。特別是幹細胞的建立,更展現了美好的前景。
