基因定位
正文
━━━━━━━━━━━━━━━━━━━━━━━━基因所屬連鎖群或染色體的測定
系譜分析法
非整倍體測交法
四分體分析法
連鎖群法
利用染色體易位的基因定位
基因在染色體上的位置的測定
根據重組頻率的基因定位
根據所測基因在某一已知染色體區段中是
否存在的基因定位
根據並發事件的基因定位
根據基因行為的定位
測定絕對位置的基因定位
基因精細結構分析
重組頻率定位法
缺失定位法
共轉導定位法
體細胞重組定位法
基因轉變的梯度定位法
小結
━━━━━━━━━━━━━━━━━━━━━━━━
基因所屬連鎖群或染色體以及基因在染色體上的位置的測定。它是遺傳學研究中的一項基本工作;至於一個基因內部的突變位點的測定則一般稱為基因精細結構分析。染色體基因定位方法多數也適用於染色體外遺傳研究中的基因定位。
基因定位是遺傳學研究中的重要環節。在遺傳學的早期研究中並未發現果蠅等生物的基因在染色體上的位置和生理功能有什麼關係。但以後發現一些有類似表型效應的基因是緊密連鎖的。例如1945年E.B.劉易斯在果蠅中發現與中胸發育有關的幾個基因相鄰接,構成一個複合座位或稱基因複合體或擬等位基因系列;1960年J.莫諾和F.雅各布報導大腸桿菌的與乳糖發酵有關的幾個基因緊密連鎖,構成一個操縱子。可見基因的位置並不是和它們的功能完全無關的,因此基因定位有助於了解基因的功能。此外,測定了某一基因在某一染色體上的位置以後,便可以用這一基因作為所屬染色體或其一部分的標記,追蹤並研究染色體的行為。例如通過分析大腸桿菌的接合過程中各個標記基因在受體菌株中出現的先後次序,就有助於了解接合過程中染色體的行為(見細菌接合);在許多生物中根據雜交子代中各個標記基因的組合,可以研究染色體干涉、染色單體干涉和染色體畸變;在育種工作中也經常通過標記基因來識別染色體的替換。1913年C.B.布里奇斯首先在果蠅中通過 X染色體的不離開現象證實了白眼基因(white,w)是在X染色體上。同年A.H.斯特蒂文特根據兩個基因之間的距離愈遠則交換頻率愈高這一假設,首先在果蠅中進行了基因定位工作。
基因所屬連鎖群或染色體的測定
系譜分析法 在早期的人類遺傳學研究中,基因所屬染色體的測定一般都通過系譜分析進行。由於女子有兩個X染色體,男子有一個X染色體和一個Y染色體,Y染色體上不存在和X染色體相應的等位基因(也就是說對於 X連鎖基因來講,男子是半合體)。因此男性患者的X連鎖致病基因必然來自母親,以後又必定傳給女兒。這種遺傳方式稱為交叉遺傳。如果在一個家系中外祖父是某種疾病的患者,母親的表型是正常的,外孫中有半數是患者,就可以判斷有關的隱性致病基因是在 X染色體上。色盲基因便是1911年通過系譜分析最早發現的人的性連鎖基因。位置在X染色體上的性連鎖基因稱為X連鎖基因。常染色體遺傳也有它自己的系譜特點(見人類遺傳性疾病)。根據系譜分析一般只能夠判斷基因屬於性染色體或常染色體,可是不能判斷究竟屬於哪一個常染色體。
非整倍體測交法 非整倍體測交法可以用來測定基因屬於哪一個常染色體。用常染色體隱性突變型純合體(a/a)和野生型二倍體(+/+)雜交,再用子一代雜合體(a/+)和隱性親本回交,在它們的子代中表型是野生型的和表型是突變型的各占50%(見孟德爾定律)。
雜交a/a×+/+
↓
回交 a/+ × a/a
↓
回交子代 a/aa/+
突變型 野生型
比例 1 ∶1
如果常染色體隱性突變型純合體和某一染色體的野生型三體 (+/+/+)品系(見染色體畸變)雜交,子一代中的三體個體再和隱性親本回交,在它們的子代中野生型和突變型之比是5∶1而不是1∶1。
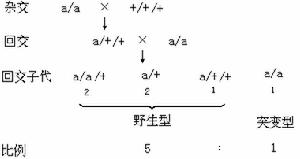
雜交 a/a× +
↓
子一代a/+∶ a
野生型突變型
比例 1 ∶ 1
根據上述三種不同的雜交結果,可見只要具備相當於每一染色體的一系列三體和單體品系,便能從雜交子代的突變型和野生型的比數中判斷任何一個突變基因所屬的染色體。小麥是多倍體植物,多倍體植物增加或減少一個染色體不會使它的生活力受到嚴重的影響,因此容易建立整套三體或單體品系,使基因定位工作得以順利進行。除了小麥等植物以外,這一方法也用在酵母菌的遺傳學研究中。
四分體分析法 由於子囊菌減數分裂所形成的四分體包被在一個子囊里,所以判斷兩個基因是否連鎖,只需計算出各種類型的四分體數(即子囊數)。如果其中一個基因所屬的連鎖群已經知道,便很容易測定另一基因是否屬於同一連鎖群。
四分體有三種,即親代二型(PD)、非親代二型(NPD)和四型 (T)。如果有關的兩個基因是連鎖的,即PD是不交換或二線雙交換的結果,NPD是四線雙交換的結果,T是單交換或三線雙交換的結果(見連鎖和交換)。如果有關的兩個基因是不連鎖的,那么雙基因雜交子代中所出現的四分體類型要看這兩個基因各自和著絲粒之間是否發生交換而定(表1)。
 基因定位
基因定位連鎖群法 利用近著絲粒距離基因的定位法 如果某一染色體上有一個離著絲粒距離較近的已知基因,另外有一個基因同樣離著絲粒很近,可是不知道它是否屬於同一染色體。把這樣兩個突變型品系進行雜交,如果這兩個基因屬於同一染色體,它們之間的重組頻率不應超過兩者的著絲粒距離之和;如果它們不屬於同一染色體,那么它們的重組頻率應是50%(見連鎖和交換)。由於這兩個基因與著絲粒距離都是較近的,所以增加了這一判斷的可靠性。用這一方法曾測得粗糙脈孢菌的連鎖群數是7。
同線法 如果一個細胞得到或丟失一條染色體則將同時得到或失去這條染色體上的全部基因。如果其中某些基因是已知的,而另一連鎖關係未知的基因恰恰和上述基因同時得到或失去,便可以判定後一基因和前一基因屬於同一連鎖群(表2)。
 基因定位
基因定位假定有五個人-鼠融合細胞株 A、B、C、D、E,它們分別保留有人染色體1、2、3中的這一個或那一個,例如細胞株A保留染色體2,細胞株D保留1、2、3這三個染色體等。分別測定甲、乙、丙、丁四種表型,如某種酶的活性或某種抗原的存在與否等。已經知道這些特性都是小鼠細胞所沒有的,從表2的結果可以看到任何一個細胞株只要測得(或不能測得)甲這一性狀時必然同時可以測得(或不能測得)丙這一性狀。這一結果說明這兩種酶或抗原蛋白的基因屬於同一染色體。其次可以看到任何一個細胞株凡是保留染色體 2的必然出現上述性狀,可見上述兩個基因都在人的2號染色體上。
單元化定位法 在構窠麴黴這一類真菌的準性生殖過程中,雜合二倍體細胞在有絲分裂時常隨機地丟失它的染色體。染色體在多次有絲分裂過程中逐條丟失而使二倍體細胞終於轉變為單倍體細胞的過程稱為單元化。如果一對染色體中帶有顯性的野生型基因的染色體丟失了,那么同源染色體上隱性基因的性狀便得以表現。此外,通過體細胞交換也可以從雜合二倍體得到表現隱性性狀的子代細胞。不過一次體細胞交換隻能導致著絲粒一側的隱性基因的性狀得以表現,而同一染色體同時發生兩次交換的機率極低,因此假如一個染色體的著絲粒兩側的隱性基因的性狀都在一個子代細胞中表現,那就說明這是帶有顯性基因的染色體丟失的結果。另一個待測的隱性突變型如果同時得以表現,這就是它和已知基因有連鎖關係的證據。
利用染色體易位的基因定位 直接觀察法 易位(見染色體畸變)使染色體上的基因改變連鎖關係,所以易位可以用來進行基因定位。如果易位所涉及的染色體是可以被識別的,那就更有利於定位工作。如果在遺傳學分析中發現某兩個連鎖群的連鎖關係都發生了改變,同時在顯微鏡下又可以辨認出有兩個染色體發生了相互易位,那么就可以知道兩個連鎖群和兩個染色體的對應關係。例如遺傳學分析的結果說明小鼠品系 T1380的相互易位涉及連鎖群LGⅡ和LGⅨ。品系RB163H的相互易位涉及連鎖群LGⅡ和LGⅫ。細胞學觀察說明前者涉及染色體 9和17,後者涉及染色體9和19,因此知道連鎖群 LGⅡ屬於染色體9,連鎖群LGⅨ屬於染色體17,連鎖群 LGⅫ屬於染色體19。
某些生物的染色體具有天然的標記,例如果蠅的唾腺染色體具有容易辨認的橫紋,玉米的染色體有容易辨認的巨大的染色粒,所以通過直接的細胞學觀察就可以辨認出易位所涉及的染色體,但是對於染色體數較多而又沒有天然標記的生物,就需用顯帶技術鑑定染色體的易位。上述小鼠的連鎖群所屬的染色體便是套用顯帶技術鑑定的(見核型)。
假連鎖法 相互易位雜合體只有在減數分裂過程中通過互動離開所形成的平衡配子才能夠存活,並使非同源染色體上的基因顯示假連鎖現象(見染色體畸變)。所以把帶有屬於已知染色體的標記基因的相互易位品系作為測交品系和一個突變型品系雜交,如果發現這一突變基因經常和標記基因的野生型等位基因相連鎖,就可以判定突變基因一定在相互易位的兩個染色體中的一個上面。例如在粗糙脈孢菌中有一個品系(圖1),它的第 Ⅰ染色體上帶有白色分生孢子基因(albino,al),第Ⅳ 染色體上帶有溫度敏感的蔓延菌落基因(colonial temperature sensitive,cot),第Ⅵ染色體上帶有黃色分生孢子基因(yellow conidia,ylo),同時它還是染色體Ⅰ-Ⅱ、Ⅳ-Ⅴ和Ⅲ-Ⅵ相互易位品系。用它作為測試菌株和任何一個突變型菌株進行雜交,就可以通過一次雜交大致上測得這一突變型所屬的染色體。例如具有這一突變性狀的雜交子代菌株總是菌絲蔓延的,就知道它的突變基因屬於染色體Ⅳ或Ⅴ。又假如這個突變型和al、cot和ylo都沒有連鎖關係,那么由於粗糙脈孢菌只有7個染色體,就可以推測這一突變基因在染色體Ⅶ上。
 基因定位
基因定位基因在染色體上的位置的測定
根據重組頻率的基因定位 同一染色體上兩個基因之間的距離愈遠,則發生交換的機會愈多,雜交子代中重組體也就愈多。所以測定雜交子代中重組體的多少,就可以知道有關的基因的距離,這是最基本的基因定位方法。A.H.斯特蒂文特把雜交子代中出現 1%重組體的兩個基因之間的距離定為一個圖距單位,後來又有人稱之為一個分摩,用來紀念首先提出交換概念的T.H.摩爾根。同一原理也適用於單倍體微生物的基因定位。
三點測驗法 在包括兩對基因的雜交中,一次雜交可以測定兩個基因之間的距離,通過三次雜交便可以測定三個基因的排列順序和距離。但是在包括三對基因的一次雜交中,便可以測定三個基因的排列順序和距離,這就是1913年由斯特蒂文特首創的三點測驗方法。例如黑腹果蠅的X染色體上有黃體基因(yellow body,y;野生型灰體,y+)、白眼基因(white eye, w;野生型紅眼,w+)和短翅基因 (miniature wing,m;野生型長翅,m+)。將黃體、白眼、短翅雌蠅
 和野生型雄蠅 (y+w+m+ 即+++)雜交,將得到的雌性雜合體
和野生型雄蠅 (y+w+m+ 即+++)雜交,將得到的雌性雜合體  再和雄性子代ywm雜交,得到子二代個體(表3)。
再和雄性子代ywm雜交,得到子二代個體(表3)。 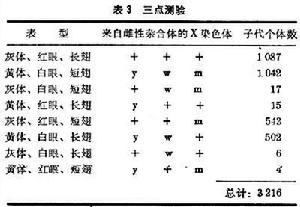 基因定位
基因定位基因y和w之間的重組頻率 =1.3%
=1.3%
基因w和m之間的重組頻率 =32.8%
=32.8%
基因y和m之間的重組頻率
 基因定位
基因定位子囊的排列方式有 6種,AAaa和aaAA這兩種稱為第一次分裂分離,AaAa、aAaA、AaaA、aAAa這四種稱為第二次分裂分離。前者基因A(a)和著絲粒之間沒有發生交換,後者A(a)和著絲粒之間發生了交換。
所以某一基因和著絲粒之間交換頻率愈高,第二次分裂分離子囊愈多。由於每次交換導致半數染色單體成為重組類型,所以

體細胞交換法 三點測驗和著絲粒距離法中所測定的都是發生在減數分裂中的染色體交換。1936年美國遺傳學家C.斯特恩在果蠅中發現體細胞在有絲分裂過程中也可以發生染色體交換(見連鎖和交換)。
50年代中G.蓬泰科爾沃等在研究構窠麴黴時發展起來一種利用體細胞交換的系統的基因定位方法。在進行有絲分裂的雜合二倍體細胞中,體細胞交換會導致在子代體細胞中出現隱性基因的純合體,這一過程稱為純合化。
如果某一個二倍體細胞的某一染色體臂上有若干個基因都呈雜合狀態,那么就可根據子代體細胞各個基因純合化的頻率推知它們的相對位置。交換隻使比交換位置更遠離著絲粒的隱性基因純合化,所以某個基因純合化的頻率愈高,它離著絲粒的距離就愈遠(圖3)。
 基因定位
基因定位根據所測基因在某一已知染色體區段中是否存在的基因定位 如果染色體的某一區段的位置是已知的,而且測得某一基因的位置在這一區段中,那么這一基因的位置也就被測定了。
缺失定位法 一個細胞中的兩個同源染色體中的一個上有一個突變基因,另一染色體上有一小段已知範圍的缺失,如果這一突變基因的位置在缺失範圍內,便不可能通過重組而得到野生型重組體;如果突變基因不在缺失範圍內,那么就可以得到野生型重組體。利用一系列已知缺失位置和範圍的缺失突變型,便能測定突變型基因的位置。
標記獲救法 這是一種結合物理圖譜製作和遺傳學分析的基因定位方法,它適用於病毒等基因組較小的生物。以大腸桿菌噬菌體ΦX174為例,把野生型噬菌體的雙鏈複製型DNA分子用限制性內切酶HindⅡ切為13個片段,把每種片段和突變型 amg的DNA單鏈在使DNA分子變性並復性的條件下混合保溫,然後用各個樣品分別轉化受體細菌。如果在某一樣品處理後的受體細菌中出現了大量的野生型噬菌體,於是就說明這一樣品中的HindⅡ片段包含著amg的相應的野生型基因,由於13個HindⅡ片段的位置在物理圖譜中全部都是已知的,因此便可以推知amg基因在染色體上的相應位置。用這一方法在ΦX174的環狀的染色體圖上已經測定了至少19個基因的位置。
根據並發事件的基因定位 位置鄰近的基因表現某些相關的行為,所以從這些行為可以推測基因的連鎖關係。
共轉導法 每一種轉導噬菌體有一定的大小,只能攜帶一定長度的供體細菌的 DNA。例如大腸桿菌噬菌體PI的頭部中只能包裝大約分子量為5.8×107的DNA,大腸桿菌的染色體DNA的分子量是2.5×109,所以PI所能包裝的 DNA至多相當於大腸桿菌的遺傳學圖上相距兩分鐘這樣一段DNA分子。如果兩個基因能同時被轉導,這兩個基因之間的距離必然較近,而且距離愈近則共轉導頻率愈高,因此可以由共轉導的頻率來推算基因間的距離。

共缺失法 缺失帶來和基因突變相同的表型。由一次缺失所造成的突變只涉及相鄰接的基因,因此可以從缺失所帶來的基因突變的分析來測定一些基因的相對位置,這一方法被廣泛套用於酵母菌的線粒體基因的定位(見染色體外遺傳)。
根據基因行為的定位 基因的某些行為可以反映它們的位置。在細菌接合過程中“雄性”細菌的染色體基因按先後順序轉移到“雌性”細菌中。一些基因組較小的病毒,整個基因組往往作為一個單位轉錄。因此接合過程中基因轉移的先後、轉錄過程中轉錄的先後或DNA複製的先後都可以在某些特殊的生物中用來作為基因定位的手段。
中斷雜交法 見細菌接合。
轉錄定位法 許多 RNA病毒的整個基因組往往作為一個單位轉錄。隨著轉錄的進行,由基因組上各個基因所編碼的蛋白質也依序在寄主細胞中出現。當寄主細胞被紫外線照射使本身的蛋白質合成受到抑制時,病毒蛋白的出現更為明顯。紫外線照射也起著抑制病毒基因組的轉錄的作用。紫外線在 RNA分子的某一部位造成損傷後,損傷的部位和它後面的基因的轉錄都將受到影響,損傷部位以前的基因的轉錄則不受影響。因為轉錄沿負鏈RNA的3′端向5′端進行,所以愈是接近3′端的基因的轉錄和由它編碼的蛋白質在寄主細胞中的合成受到紫外線損傷的影響愈小,而愈是接近 5′端的基因和相應的蛋白質的合成愈容易為紫外線照射所抑制。因此只要先用相同劑量的紫外線照射待測病毒,然後再測出寄主細胞中該病毒編碼的各種蛋白質的產量,便可以推知該病毒各個基因的位置。
測定絕對位置的基因定位 細胞學圖 通過種種方法可以測得基因之間的距離,但圖距並不表示絕對長度,而且在不同的生物中同一圖距代表不同的實際長度。通過細胞遺傳學的方法可以測定基因的實際位置,這樣繪製的基因位置圖稱為細胞學圖,而通過一般遺傳學方法繪製的圖則稱為遺傳學圖。
在雜合的二倍體生物中,由於顯性的野生型基因的存在,隱性的突變基因得不到表現。如果帶有野生型基因的這一染色體發生了一個缺失,而缺失部分又正好包括這一野生型基因,那么同源染色體上相應的隱性基因的突變性狀便得以表現(見染色體畸變)。果蠅具有便於觀察染色體細微結構的唾腺染色體,它上面的橫紋缺失可以在光學顯微鏡下識別。通過一系列的雜交,可以得到某一隱性突變基因和一系列的缺失染色體組合在一起的果蠅,對於這些雜合體果蠅進行染色體分析和性狀觀察,便可以判斷某隱性突變基因在染色體上的真實位置。
從果蠅的X染色體上包括白色複眼(white eye,w)和小糙眼(facet,fa)區域的分析結果(圖4)可以看到凡是缺失3C7這一橫紋的雜合體都呈現小糙眼突變型性狀,說明fa基因位置在唾腺染色體的3C7橫紋處。
 基因定位
基因定位各種限制性核酸內切酶具有各自的識別順序。這些識別順序可以作為DNA部位的標記,用不同的限制酶處理同一DNA分子,通過對酶切產生的DNA片段的大小和位置的分析,可以繪製出某一 DNA分子的限制圖譜。此外,每一個DNA分子上富含A∶T鹼基對和富含 G嗈C鹼基對的區域的分布各不相同。富含A∶T鹼基對的區域比富含G嗈C鹼基對的區域更易變性。所以在嚴格控制的變性條件下每一種 DNA分子具有變性環的特定分布形式,構成部分變性圖。
分子雜交法 分子雜交和體細胞遺傳學相結合的方法也可以用來測定人的基因的絕對位置。用體細胞遺傳學方法,可以得到只含有某一條人類染色體的人-倉鼠雜種細胞的克隆。然後可以進一步取得這一人類染色體發生各種缺失的克隆。把從這一系列缺失克隆中提取出來的 DNA吸附在硝酸纖維素濾膜上。再把人的基因文庫中的各個基因的 DNA片段用32P標記製成探針,然後用探針分別與膜上吸附的 DNA進行分子雜交測驗,能雜交者表示它的缺失部分不包括這一基因。再結合染色體顯帶技術便可以測定這一基因在染色體上的絕對位置。缺失克隆的數目愈多,測定的位置就愈精確。
基因精細結構分析
重組頻率定位法 原理和在高等動植物中用雜交子代中重組頻率的高低來計算兩個基因間的距離沒有不同。不過在微生物中一個菌落或一個噬菌斑代表一個個體,因而便於通過大量的雜交子代的觀察來進行精細結構分析;而且往往採用選擇性培養方法淘汰沒有發生重組的親本,使分析的效率和精密度進一步提高。不過精細結構的重組頻率容易受到突變位置本身的影響,這就是標記影響。
缺失定位法 原理和基因的缺失定位相同,不過需要具備種類更多而差別更為細微的缺失菌株。在大腸桿菌的 T4噬菌體中曾經獲得一系列快速溶菌突變型rⅡ基因部分缺失突變型,利用這些突變型可以迅速測定任何一個 rⅡ點突變在rⅡ基因中的位置。圖5中的每一編號標明的橫線表示一個缺失突變型的缺失範圍。定位分兩步進行,首先測定大的範圍,然後測定精確的位置。例如有一個點突變型和缺失突變型 A105和638能發生重組而和其餘五個不發生重組,就可知它的位置在區域A5中,然後進一步利用A5中三個缺失突變型1605、1589、PB230進行更精確的定位(圖5)。可見只要有足夠多的缺失突變型,就可使位置測定得十分精確。這一方法在實際操作上很簡單,因為只要觀察有沒有噬菌斑出現,而無須計算噬菌斑的數目。
 基因定位
基因定位體細胞重組定位法 原理相同於基因純合化的定位方法。由於體細胞交換髮生得較少,所以常用 X射線處理雜合體使之發生更多的體細胞交換。
基因轉變的梯度定位法 一個基因內部的各個點突變的基因轉變常呈梯度現象,即在這基因的一端發生基因轉變的頻率最高,在另一端則最低,在兩端之間存在著一個轉變頻率的梯度。對於任何一個未知位置的點突變,可以通過基因轉變頻率的測定進行精細結構定位。這一方法的套用限於一次減數分裂產物包被在一個囊裡面的子囊菌,而且限於影響子囊孢子顏色和形狀的基因。
小 結
基因定位既是遺傳學研究的基本方法,又隨著遺傳學研究的不斷深入而出現新的方法。長期以來基因定位工作離不開雜交試驗,也離不開突變基因,但自從開展了分子遺傳學的研究以後,便出現了不必通過雜交也不依賴於突變基因的基因定位方法,例如分子雜交定位等方法。可以預見,隨著研究工作的深入,必將出現更為精確的基因定位方法。
配圖
 基因定位
基因定位