
簡介
 同工酶圖譜
同工酶圖譜分類
基因性或原級
由不同基因產生的肽鏈而衍生的同工酶。這裡所指的不同基因可以在不同染色體或在同一染色體的不同位點上,例如LDH中A、B兩條肽鏈的基因分別在第11及第12對染色體上,唾液澱粉酶和胰澱粉酶的基因在第1對染色體的不同位點上。這類同工酶因分子結構差異較大,彼此間無交叉免疫。但同工酶的不同基因也可以是同源染色體的等位基因,這種成對的等位基因上兩個基因結構不同的情況,在遺傳學上稱為雜合子。雜合子在同一個體中可合成同一種酶的兩種不同肽鏈,或亞基,這兩種亞基尚可雜交,形成同工酶。在生物群體的不同個體中,有時同一基因位點上的一個(對雜合子來說)或一對(對純合子來說)基因也可發生遺傳變異,從而產生變異的酶,出現群體中的遺傳多態。不同個體中這些遺傳變異的酶也屬於基因性同工酶。其在免疫學上常有交叉反應。
由同一基因轉錄出前體核糖核酸(前體RNA),經過不同的加工剪接過程而生成多種不同的mRNA,再轉譯出多種肽鏈,從而組成同工酶。這類同工酶因發現較晚,在國際上尚無統一命名,彼此間也有交叉免疫。
次生或轉譯後
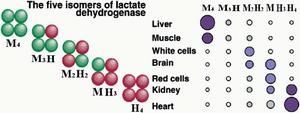 同工酶
同工酶國際生化協會命名委員會(CBN)建議只將原級同工酶列為同工酶,而將次生同工酶稱為共合酶,但不少生化學家還是把上述各類酶的不同結構形式都包括在廣義的同工酶概念中。
結構特點
同工酶是指催化相同的化學反應,而酶蛋白的分子結構、理化性質乃至免疫學性質不同的一組酶。同工酶是由不同基因或等位基因編碼的多肽鏈,或由同一基因轉錄生成的不同mRNA翻譯的不同多肽鏈組成的蛋白質。翻譯後經修飾生成的多分子形式不在同工酶之列。
同工酶存在於同一種屬或同一個體的不同組織或同一細胞的不同亞細胞結構中。如乳酸脫氫酶是四聚體酶。亞基有兩型:骨骼肌型(M型)和心肌型(H型)。同工酶的基因必須先轉錄成同工酶的信使核糖核酸(mRNA)、後者再轉譯產生組成同工酶的肽鏈,不同的肽鏈可以不聚合的單體形式存在,也可聚合成純聚體或雜交體,從而形成同一種酶的不同結構形式。
套用
 黃河鯉---同工酶
黃河鯉---同工酶在醫學方面,同工酶是研究癌瘤發生的重要手段,癌瘤組織的同工酶譜常發生胚胎化現象,即合成過多的胎兒型同工酶。如果這些變化可反映到血清中,則可利用血清同工酶譜的改變來診斷癌瘤。此外,因同工酶譜有臟器特異性,故測定血清同工酶常可較特異地反映某一臟器的病變,如血清的LDH1(B4)或MB型肌酸激酶(CK-MB)增加是診斷心肌梗死較特異的指標,較測定血清LDH或肌酸激酶(CK)總活力更為可靠。
酶相關名詞
酶 | 酶動力學 | 催化常數 | 酵素梅 | 競爭性抑制 | 非特異性抑制劑 | 酶多重性 | 多酶體系 | 開特 | 全酶 | 米氏常數 | 酶催化機制 | 催化部位 | 非競爭性抑制 | 抑制結構域 | 酶多態性 | 多酶複合物 | 活化能 | 米氏方程 | 鎖鑰學說 | 催化亞基 | 反競爭性抑制 | 協同催化 | 酶解作用 | 活性部位 | 同工酶 | 米氏動力學 | 誘導契合學說 | 不可逆抑制 | 非特異性抑制 | 協作抑制 |酶複合物| 酶活性 | 最適pH | 酶原 | 雙倒數作圖法 | 催化活性 | 可逆抑制 | 競爭性抑制劑 | 反饋抑制 | 酶系 | 酶單位 | 最適溫度
