
概述
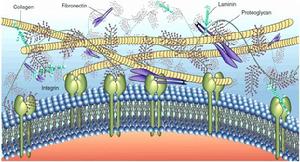 細胞外基質的成分
細胞外基質的成分細胞外基質(extracellularmatrixc,ECM)是由動物細胞合成並分泌到胞外、分布在細胞表面或細胞之間的大分子,主要是一些多糖和蛋白,或蛋白聚糖。這些物質構成複雜的網架結構,支持並連線組織結構、調節組織的發生和細胞的生理活動。細胞外基質是動物組織的一部分,不屬於任何細胞。它決定結締組織的特性,為細胞的生存及活動提供適宜的場所,並通過信號轉導系統影響細胞的形狀、代謝、功能、遷移、增殖和分化。
成分
 細胞外基質
細胞外基質細胞外基質的組成可分為三大類:①糖胺聚糖(glycosaminoglycans)、蛋白聚糖(proteoglycan),它們能夠形成水性的膠狀物,在這種膠狀物中包埋有許多其它的基質成分;②結構蛋白,如膠原和彈性蛋白,它們賦予細胞外基質一定的強度和韌性;③粘著蛋白(adhesive):如纖粘連蛋白和層粘聯蛋白,它們促使細胞同基質結合。其中以膠原和蛋白聚糖為基本骨架在細胞表面形成纖維網狀複合物,這種複合物通過纖粘連蛋白或層粘連蛋白以及其他的連線分子直接與細胞表面受體連線;或附著到受體上。由於受體多數是膜整合蛋白,並與細胞內的骨架蛋白相連,所以細胞外基質通過膜整合蛋白將細胞外與細胞內連成了一個整體。
構成細胞外基質的大分子種類繁多,可大致歸納為四大類:膠原、非膠原糖蛋白、氨基聚糖與蛋白聚糖、以及彈性蛋白。
上皮組織、肌組織及腦與脊髓中的ECM含量較少,而結締組織中ECM含量較高。細胞外基質的組分及組裝形式由所產生的細胞決定,並與組織的特殊功能需要相適應。例如,角膜的細胞外基質為透明柔軟的片層,肌腱的則堅韌如繩索。細胞外基質不僅靜態的發揮支持、連線、保水、保護等物理作用,而且動態的對細胞產生全方位影響。
膠原
膠原是動物體內含量最豐富的蛋白質,約占人體蛋白質總量的30%以上。它遍布於體內各種器官和組織,是細胞外基質中的框架結構,可由成纖維細胞、軟骨細胞、成骨細胞及某些上皮細胞合成並分泌到細胞外。
 細胞外基質
細胞外基質已發現的膠原至少有19種,由不同的結構基因編碼,具有不同的化學結構及免疫學特性。Ⅰ、Ⅱ、Ⅲ、Ⅴ及Ⅺ型膠原為有橫紋的纖維形膠原。
各型膠原都是由三條相同或不同的肽鏈形成三股螺鏇,含有三種結構:螺鏇區,非螺鏇區及球形結構域。其中Ⅰ型膠原的結構最為典型。
Ⅰ型膠原的原纖維平行排列成較粗大的束,成為光鏡下可見的膠原纖維,抗張強度超過鋼筋。其三股螺鏇由二條α1(Ⅰ)鏈及一條α2(Ⅰ)鏈構成。每條α鏈約含1050個胺基酸殘基,由重複的Gly-X-Y序列構成。X常為Pro(脯氨酸),Y常為羥脯氨酸或羥賴氨酸殘基。重複的Gly-X-Y序列使α鏈捲曲為左手螺鏇,每圈含3個胺基酸殘基。三股這樣的螺鏇再相互盤繞成右手超螺鏇,即原膠原。
 細胞外基質
細胞外基質原膠原分子間通過側向共價交聯,相互呈階梯式有序排列聚合成直徑50~200nm、長150nm至數微米的原纖維,在電鏡下可見間隔67nm的橫紋。膠原原纖維中的交聯鍵是由側向相鄰的賴氨酸或羥賴氨酸殘基氧化後所產生的兩個醛基間進行縮合而形成的。
原膠原共價交聯後成為具有抗張強度的不溶性膠原。胚胎及新生兒的膠原因缺乏分子間的交聯而易於抽提。隨年齡增長,交聯日益增多,皮膚、血管及各種組織變得僵硬,成為老化的一個重要特徵。
人α1(Ⅰ)鏈的基因含51個外顯子,因而基因轉錄後的拼接十分複雜。翻譯出的肽鏈稱為前α鏈,其兩端各具有一段不含Gly-X-Y序列的前肽。三條前α鏈的C端前肽借二硫鍵形成鏈間交聯,使三條前α鏈“對齊”排列。然後從C端向N端形成三股螺鏇結構。前肽部分則呈非螺鏇捲曲。帶有前肽的三股螺鏇膠原分子稱為前膠原(procollagen)。膠原變性後不能自然復性重新形成三股螺鏇結構,原因是成熟膠原分子的肽鏈不含前肽,故而不能再進行“對齊”排列。
前α鏈在粗面內質網上合成,並在形成三股螺鏇之前於脯氨酸及賴氨酸殘基上進行羥基化修飾,脯氨酸殘基的羥化反應是在與膜結合的脯氨醯-4羥化酶及脯氨醯-3羥化酶的催化下進行的。維生素C是這兩種酶所必需的輔助因子。維生素C缺乏導致膠原的羥化反應不能充分進行,不能形成正常的膠原原纖維,結果非羥化的前α鏈在細胞內被降解。因而,膳食中缺乏維生素C可導致血管、肌腱、皮膚變脆,易出血,稱為壞血病。
纖粘連蛋白
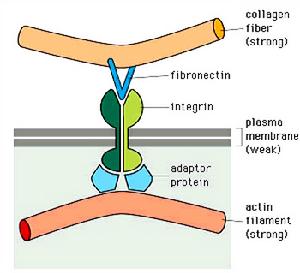 細胞外基質
細胞外基質每條FN肽鏈約含2450個胺基酸殘基,整個肽鏈由三種類型(Ⅰ、Ⅱ、Ⅲ)的模組(module)重複排列構成。具有5-7個有特定功能的結構域,由對蛋白酶敏感的肽段連線。這些結構域中有些能與其它ECM(如膠原、蛋白聚糖)結合,使細胞外基質形成網路;有些能與細胞表面的受體結合,使細胞附著與ECM上。
FN肽鏈中的一些短肽序列為細胞表面的各種FN受體識別與結合的最小結構單位。例如,在肽鏈中央的與細胞相結合的模組中存在RGD(Arg-Gly-Asp)序列,為與細胞表面某些整合素受體識別與結合的部位。化學合成的RGD三肽可抑制細胞在FN基質上粘附。
細胞表面及細胞外基質中的FN分子間通過二硫鍵相互交聯,組裝成纖維。與膠原不同,FN不能自發組裝成纖維,而是通過細胞表面受體指導下進行的,只存在於某些細胞(如成纖維細胞)表面。轉化細胞及腫瘤細胞表面的FN纖維減少或缺失系因細胞表面的FN受體異常所致。
層粘連蛋白
層粘連蛋白(laminin,LN)LN也是一種大型的糖蛋白,與Ⅳ型膠原一起構成基膜,是胚胎髮育中出現最早的細胞外基質成分。LN分子由一條重鏈(α)和二條輕鏈(β、γ)借二硫鍵交聯而成,外形呈十字形,三條短臂各由三條肽鏈的N端序列構成。每一短臂包括二個球區及二個短桿區,長臂也由桿區及球區構成。LN分子中至少存在8個與細胞結合的位點。例如,在長臂靠近球區的。鏈上有IKVAV五肽序列可與神經細胞結合,並促進神經生長。鼠LNα1鏈上的RGD序列,可與αvβ3整合素結合。
現已發現7種LN分子,8種亞單位(α1,α2,α3,β1,β2,β3,γ1,γ2),與FN不同的是,這8種亞單位分別由8個結構基因編碼。LN是含糖量很高(占15-28%)的糖蛋白,具有50條左右N連線的糖鏈,是迄今所知糖鏈結構最複雜的糖蛋白。而且LN的多種受體是識別與結合其糖鏈結構的。基膜是上皮細胞下方一層柔軟的特化的細胞外基質,也存在於肌肉、脂肪和許旺細胞(schwanncell)周圍。它不僅僅起保護和過濾作用,還決定細胞的極性,影響細胞的代謝、存活、遷移、增殖和分化。
基膜中除LN和Ⅳ型膠原外,還具有entactin、perlecan、decorin等多種蛋白,其中LN與entactin(alsocallednidogen)形成1:1緊密結合的複合物,通過nidogen與Ⅳ型膠原結合。
氨基聚糖與蛋白聚糖
1.氨基聚糖(glycosaminoglycan,GAG)
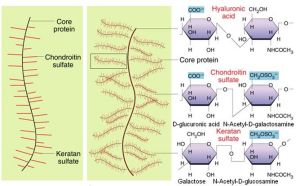
GAG是由重複二糖單位構成的無分枝長鏈多糖。其二糖單位通常由氨基已糖(氨基葡萄糖或氨基半乳糖)和糖醛酸組成,但硫酸角質素中糖醛酸由半乳糖代替。氨基聚糖依組成糖基、連線方式、硫酸化程度及位置的不同可分為六種,即:透明質酸、硫酸軟骨素、硫酸皮膚素、硫酸乙醯肝素、肝素、硫酸角質素。
透明質酸(hyaluronicacid,HA)是唯一不發生硫酸化的氨基聚糖,其糖鏈特別長。氨基聚糖一般由不到300個單糖基組成,而HA可含10萬個糖基。在溶液中HA分子呈無規則捲曲狀態。如果強行伸長,其分子長度可達20μm。HA整個分子全部由葡萄糖醛酸及乙醯氨基葡萄糖二糖單位重複排列構成。由於HA分子表面有大量帶負電荷的親水性基團,可結合大量水分子,因而即使濃度很低也能形成粘稠的膠體,占據很大的空間,產生膨壓。
細胞表面的HA受體為CD44及其同源分子,屬於hyaladherin族。所有能結合HA的分子都具相似的結構域。HA雖不與蛋白質共價結合,但可與許多種蛋白聚糖的核心蛋白質及連線蛋白質借非共價鍵結合而參加蛋白聚糖多聚體的構成,在軟骨基質中尤其如此。除HA及肝素外,其他幾種氨基聚糖均不游離存在,而與核心蛋白質共價結合構成蛋白聚糖。
2.蛋白聚糖(proteoglycan)
蛋白聚糖是氨基聚糖(除透明質酸外)與核心蛋白質(coreprotein)的共價結合物。核心蛋白質的絲氨酸殘基(常有Ser-Gly-X-Gly序列)可在高爾基複合體中裝配上氨基聚糖(GAG)鏈。其糖基化過程為通過逐個轉移糖基首先合成由四糖組成的連線橋(Xyl-Gal-Gal-GlcUA),然後再延長糖鏈,並對所合成的重複二糖單位進行硫酸化及差向異構化修飾。一個核心蛋白質分子上可以連線1至100個以上GAG鏈。與一個核心蛋白質分子相連的GAG鏈可以是同種或不同種的。
許多蛋白聚糖單體常以非共價鍵與透明質酸形成多聚體。核心蛋白質的N端序列與CD44分子結合透明質酸的結構域具有同源性,故亦屬hyaladherin族。蛋白聚糖多聚體的分子量可達108KD以上。其體積可超過細菌。如構成軟骨的Aggrecan,其GAG主要是硫酸軟骨素(chondroitinsulfate,CS),但還有硫酸角質素(keratansulfate,KS)。其含量不足或代謝障礙可引起長骨發育不良,四肢短小。
彈性蛋白
彈性蛋白纖維網路賦予組織以彈性,彈性纖維的伸展性比同樣橫截面積的橡皮條至少大5倍。
彈性蛋白由二種類型短肽段交替排列構成。一種是疏水短肽賦予分子以彈性;另一種短肽為富丙氨酸及賴氨酸殘基的α螺鏇,負責在相鄰分子間形成交聯。彈性蛋白的胺基酸組成似膠原,也富於甘氨酸及脯氨酸,但很少含羥脯氨酸,不含羥賴氨酸,沒有膠原特有的Gly-X-Y序列,故不形成規則的三股螺鏇結構。彈性蛋白分子間的交聯比膠原更複雜。通過賴氨酸殘基參與的交聯形成富於彈性的網狀結構。
在彈性蛋白的外圍包繞著一層由微原纖維構成的殼。微原纖維是由一些糖蛋白構成的。其中一種較大的糖蛋白是fibrillin,為保持彈性纖維的完整性所必需。在發育中的彈性組織內,糖蛋白微原纖維常先於彈性蛋白出現,似乎是彈性蛋白附著的框架,對於彈性蛋白分子組裝成彈性纖維具有組織作用。老年組織中彈性蛋白的生成減少,降解增強,以致組織失去彈性。
生物學作用
細胞外基質不只具有連線、支持、保水、抗壓及保護等物理學作用,而且對細胞的基本生命活動發揮全方位的生物學作用。鑒於細胞外間質的多樣性,細胞外間質有多方面的功能。例如,為細胞提供支持和固定、提供組織間的分離方法、調節細胞間的溝通。細胞外間質調節細胞的動態行為。此外細胞外間質還吸收了多種細胞生長因子和蛋白酶。當生理條件變化時,激活蛋白酶而釋放這些細胞因子,而無需從頭合成這些因子,從而迅速激活細胞功能。
1.影響細胞的存活、生長與死亡
 細胞外基質
細胞外基質正常真核細胞,除成熟血細胞外,大多須粘附於特定的細胞外基質上才能抑制凋亡而存活,稱為定著依賴性(anchoragedependence)。例如,上皮細胞及內皮細胞一旦脫離了細胞外基質則會發生程式性死亡。此現象稱為凋亡(anoikis,aGreekwordmeaning“homelessness”)。
不同的細胞外基質對細胞增殖的影響不同。例如,成纖維細胞在纖粘連蛋白基質上增殖加快,在層粘連蛋白基質上增殖減慢;而上皮細胞對纖粘連蛋白及層粘連蛋白的增殖反應則相反。腫瘤細胞的增殖喪失了定著依賴性,可在半懸浮狀態增殖。
2.決定細胞的形狀
體外實驗證明,各種細胞脫離了細胞外基質呈單個游離狀態時多呈球形。同一種細胞在不同的細胞外基質上粘附時可表現出完全不同的形狀。上皮細胞粘附於基膜上才能顯現出其極性。細胞外基質決定細胞的形狀這一作用是通過其受體影響細胞骨架的組裝而實現的。不同細胞具有不同的細胞外基質,介導的細胞骨架組裝的狀況不同,從而表現出不同的形狀。
3.控制細胞的分化
細胞通過與特定的細胞外基質成分作用而發生分化。例如,成肌細胞在纖粘連蛋白上增殖並保持未分化的表型;而在層粘連蛋白上則停止增殖,進行分化,融合為肌管。
4.參與細胞的遷移
細胞外基質可以控制細胞遷移的速度與方向,並為細胞遷移提供“腳手架”。例如,纖粘連蛋白可促進成纖維細胞及角膜上皮細胞的遷移;層粘連蛋白可促進多種腫瘤細胞的遷移。細胞的趨化性與趨觸性遷移皆依賴於細胞外基質。這在胚胎髮育及創傷癒合中具有重要意義。細胞的遷移依賴於細胞的粘附與細胞骨架的組裝。細胞粘附於一定的細胞外基質時誘導粘著斑的形成,粘著斑是聯繫細胞外基質與細胞骨架“鉚釘”。
由於細胞外基質對細胞的形狀、結構、功能、存活、增殖、分化、遷移等一切生命現象具有全面的影響,因而無論在胚胎髮育的形態發生、器官形成過程中,或在維持成體結構與功能完善(包括免疫應答及創傷修復等)的一切生理活動中均具有不可忽視的重要作用。
腎臟作用
1、細胞外基質的作用
1、細胞外基質(ECM)是構成腎臟組織結構框架的重要膠原,發現共有18種屬於ECM膠原,其中主要有Ⅰ、Ⅲ、Ⅴ、Ⅶ膠原,Ⅳ而為基底膜膠原;
2、ECM是組織生長和受損後修復的重要物質;
3、ECM的蛋白分子可通過與阻止細胞表面的粘附因子的結合對阻止細胞的趨化、增生、分化以及細胞因子的合成與分泌,起著重要的調節作用。
4、ECM擔負著重建細胞功能和維持細胞體內平衡的作用。
2、ECM與腎臟纖維化
各種原發性和/或繼發性致病原因所導致ECM的合成異常增加、降解減少,促使大量ECM積聚而沉積於腎小球、腎間質內,導致腎臟各級血管堵塞,混亂分隔形成腎臟組織形態學改變,最終導致腎單位喪失,腎功能衰竭,進一步發展成為不可逆轉的腎單位硬化。
醫學作用
目前學者們一致認為惡性腫瘤的侵蝕、轉移是一個動態的、連續的過程。腫瘤細胞首先從原發部位脫落,侵入到細胞外基質(extra
 細胞外基質
細胞外基質ECM由BM和細胞間質組成,為腫瘤轉移的重要組織屏障。腫瘤細胞通過其表面受體與ECM中的各種成分粘附後激活或分泌蛋白降解酶類來降解基質,從而形成局部溶解區,構成了腫瘤細胞轉移運行通道。一般惡性程度高的腫瘤細胞具有較強的蛋白水解作用,可侵蝕破壞包膜,促進轉移。較為關注的酶主要是絲氨酸蛋白酶類,如纖溶酶原激活物(plasminogenactivator,PA)和金屬蛋白酶(metalproteinase,MP)類,如膠原酶IV、基質降解酶、透明質酸酶。
惡性腫瘤的發生、發展、侵襲和轉移常常伴有細胞外基質(extracellularmatrix,ECM)及其細胞表面受體表達的變化。正常肝細胞沒有基膜,也不表達層粘連蛋白(laminin,LN)的特異性整合素族受體α6β1;而在肝細胞癌(humanhepatocellularcarcinoma,HCC)組織中,LN和α6β1不僅表達水平升高,呈明顯的共分布,而且其高水平表達與肝癌患者的預後呈負相關,提示HCC細胞可能通過α6β1受體接受來自LN的信號,從而對肝癌細胞的侵襲行為起著不可忽視的作用。肝癌的發病過程中往往早期就出現門靜脈侵襲、肝內轉移以及肝外肺臟和骨組織的轉移,肝癌的侵襲、轉移和術後復發是影響患者預後的主要因素。基質金屬蛋白酶(matrixmeta-lloproteinases,MMPs)對ECM的降解是腫瘤細胞侵襲和轉移的關鍵環節之一,多種惡性腫瘤都伴有MMPs分泌水平和活性的增高。
研究
2012年9月,紐約時報報導稱,匹茲堡大學醫學院的外科醫生彼得·魯賓帶領的研究人員從動物組織上取得細胞外基質,通過它成功讓人體的某些器官或肌肉重生,且不會遭到人體免疫系統的排斥。
