![染色體[10]](/img/3/b1c/nBnauM3XyIjN3YTMwEDN2UTO1UTM1QDN5MjM5ADMwAjMwUzLxQzL2MzLt92YucmbvRWdo5Cd0FmLzE2LvoDc0RHa.jpg "染色體[10]")
簡介
 染色體
染色體染色體是細胞核中載有遺傳信息(基因)的物質,在顯微鏡下呈絲狀或棒狀,由核酸和蛋白質組成,在細胞發生有絲分裂時期容易被鹼性染料著色 ,因此而得名。在無性繁殖物種中,生物體內所有細胞的染色體數目都一樣。而在有性繁殖物種中,生物體的體細胞染色體成對分布,稱為二倍體。性細胞如精子、卵子等是單倍體,染色體數目只是體細胞的一半。
哺乳動物雄性個體細胞的性染色體對為XY;雌性則為XX。鳥類的性染色體與哺乳動物不同:雄性個體的是ZZ,雌性個體為ZW。
 染色體
染色體研究歷史
 染色體
染色體1879年,由德國生物學家弗萊明(altherFlemming,1843~1905年) 經過實驗發現。
1883年,魯克斯(W·Roux)觀察到細胞核內能夠被染色的絲狀體。
1888年,德國人沃爾德耶(W·Waldeyer)稱這種絲狀體為染色體(英文:chromosome;希臘文:chroma=顏色,soma=體),意即可染色的小體,並猜測染色體與遺傳有關。
1902年,美國生物學家薩頓和鮑維里通過觀察細胞的減數分裂時又發現染色體是成對的,並推測基因位於染色體上。
1928年摩爾根證實了染色體是遺傳基因的載體,從而獲得了生理醫學諾貝爾獎。
1956年莊有興等人明確了人類每個細胞有46條染色體,46條染色體按其大小、形態配成23對,第一對到第二十二對叫做常染色體,為男女共有,第二十三對是一對性染色體。
 染色體
染色體1900年麥克朗(McClung,C.E)等就發現了決定性別的染色體。他們採用的材料多為蚱蜢和其它直翅目昆蟲。
1902年麥克朗發現了一種特殊的染色體,稱為副染色體(accessory chromosome)。在受精時,它決定昆蟲的性別。
1906年威爾遜(Wilson,E.B)觀察到另一種半翅目昆蟲(Proteror)的雌體有6對染色體,而雄性只有5對,另外加一條不配對的染色體,威爾遜稱其為X染色體,其實雌性是有一對性染色體,雄性為XO型。
在1905年斯蒂文斯(Stevens,N)發現擬步行蟲屬(Tenebriomolitor)中的一種甲蟲雌雄個體的染色體數目是相同的,但在雄性中有一對是異源的,大小不同,其中有一條雌性中也有,但是是成對的;另一條雌性中怎么也找不到,斯蒂文斯就稱之為Y染色體。在黑腹果蠅中也發現了相同的情況,果蠅共有4對染色體,在雄性中有一對是異形的染色體。
在1914年塞勒(Seiler,J)證明了在雄蛾中染色體都是同形的,而在雌蛾中有一對異形染色體。
2005年3月17日,在《Nature》雜誌上發表的一篇文章宣告基本完成對人類X染色體的全面分析。
類型
在有不同性別的生物體內,有兩個基本類型的染色體:性染色體和常染色體。前者控制性聯遺傳特徵,後者控制著除性聯遺傳特徵以外的全部遺傳特徵。人體共有22對常染色體和一對性染色體。男女的性染色體不同,男性由一個X性染色體和一個Y性染色體組成,而女性則有兩個X性染色體。第22對染色體是常染色體中最後一對,形體較小,但它與免疫系統、先天性心臟病、精神分裂、智力遲鈍和白血病以及多種癌症相關。
形態
染色體在細胞分裂之前才形成。在細胞的代謝期或間期,染色體分散成一級結構或伸展開的脫氧核糖核酸分子,組成細胞核內的染色質或核質。
染色體的形態以中期時最為典型。每條染色體由兩條染色單體組成,中間狹窄處稱為著絲點(centromere),又稱主縊痕,它將染色體分為短臂(p)和長臂(q)。按著絲粒位置的不同,人類染色體可分為中著絲粒染色體、亞中著絲粒染色體和近端著絲粒染色體等3種類型。近端著絲粒染色體的短臂末端有一個叫做隨體的結構,它呈圓球形,中間以細絲與短臂相連。有的染色體長臂上還可看到另一些較小的狹窄區,稱為次縊痕。染色體臂的末端存在著一種叫做端粒(telomere)的結構,它有保持染色體完整性的功能。
不同狀態
基因和染色體
基因在細胞里並非一盤“散沙”或“散兵游勇”,它們大多有規律地集中在細胞核內的染色體地(chromosome)上,而且每一種生物細胞內染色體的形態和數目都是一定的。染色體複製時
染色體在複製以後,含有縱向並列的兩個染色單體(chromatids),只有在著絲粒(centromere)區域仍聯在一起。著絲粒在染色體上的位置是固定的。由於著絲粒位置的不同,把染色體分成大致相等或長短不等的兩臂(arms)。著絲粒的位置在染色體中間或中間附近時,染色體兩臂的長度差不多,這著絲粒叫做中間著絲粒或亞中間著絲粒。著絲粒的位置靠近染色體的一端時,根據著絲粒離開端部的遠近,這著絲粒叫做近端部著絲粒或端部著絲粒。著絲粒所在的地方往往表現為一個縊痕,所以著絲粒又稱初級縊痕(primaryconstriction)。有些染色體上除了初級縊痕以外,還有一個次級縊痕(secondaryconstriction),連上一個叫做隨體(satellite)的遠端染色體小段。次級縊痕的位置也是固定的。在細胞分裂將結束時,核內出現一個到幾個核仁,核仁總是出現在次級縊痕的地方,所以次級縊痕也叫做核仁形成區(nucleolarorganizer)如圖:著絲粒是有絲分裂或減數分裂中的染色體高度壓縮的一個區域,在此紡錘體纖維與其結合。複雜的DNA序列構成了著絲粉。發育的麵包酵母(啤酒酵母)的著絲粒長度約為220個鹼基對,並且通過多種與剩餘DNA結合組蛋白有明顯區別的蛋白質共同保護其免受限制性核酸內切酶(簡稱限制酶)的消化。雖然著絲粒區域通過特殊的蛋自質保護其免受限制性內切核酸酶的攻擊,但該區域投有核小體而且被去凝聚,這似乎說明了在有絲分裂和減數分裂過程中著絲粒區域被高度縮窄原因。著絲粒的220對鹼基序列兩側是限制性內切核酸酶敏感位點,該位點的功能也許是促進DNA的斷裂,有助於染色單體在後期的相互分離。限制性內切核酸酶是一種在核酸內特殊位點進行切割的酶類。
結構
 染色體
染色體核小體
核小體是染色體結構的最基本單位。核小體的核心是由4種組蛋白(H2A、H2B、H3和H4)各兩個分子構成的扁球狀8聚體。
脫氧核糖核酸分子具有典型的雙螺旋結構,一個脫氧核糖核酸分子就像是一條長長的雙螺旋的飄帶。一條染色體有一個脫氧核糖核酸分子。脫氧核糖核酸雙螺旋依次在每個組蛋白8聚體分子的表面盤繞約1.75圈,其長度相當於140個鹼基對。組蛋白8聚體與其表面上盤繞的脫氧核糖核酸分子共同構成核小體。在相鄰的兩個核小體之間,有長約50~60個鹼基對的脫氧核糖核酸連線線。在相鄰的連線線之間結合著一個第5種組蛋白(H1)的分子。密集成串的核小體形成了核質中的100埃左右的纖維,這就是染色體的“一級結構”。在這裡,脫氧核糖核酸分子大約被壓縮了7倍。
螺線體
染色體的一級結構經螺旋化形成中空的線狀體,稱為螺線體或核絲,這是染色體的“二級結構”,其外徑約300埃,內徑100埃,相鄰螺旋間距為110埃。螺絲體的每一周螺旋包括6個核小體,因此脫氧核糖核酸的長度在這個等級上又被再壓縮了6倍。
超螺旋體
300埃左右的螺線體(二級結構)再進一步螺旋化,形成直徑為0.4微米(μm)的筒狀體,稱為超螺旋體。這就是染色體的“三級結構”。到這裡,脫氧核糖核酸又再被壓縮了40倍。
四級結構
超螺旋體進一步摺疊盤繞後,形成染色單體—染色體的“四級結構”。兩條染色單體組成一條染色體。到這裡,脫氧核糖核酸的長度又再被壓縮了5倍。從染色體的一級結構到四級結構,脫氧核糖核酸分子一共被壓縮了7×6×40×5=8400倍。例如,人的染色體中脫氧核糖核酸分子伸展開來的長度平均約為幾個厘米,而染色體被壓縮到只有幾個微米長。
現代關於染色體超微結構的概念
 電子顯微鏡下的人類染色體
電子顯微鏡下的人類染色體電子顯微鏡下的人類染色體。染色體的超微結構顯示染色體是由直徑僅100埃(Å)的脫氧核糖核酸-組蛋白高度螺旋化的纖維所組成。每一條染色單體可看作一條雙螺旋的脫氧核糖核酸分子。有絲分裂間期時,解螺旋而形成無限伸展的細絲,此時不易為染料所著色,光鏡下呈無定形物質,稱之為染色質。有絲分裂時脫氧核糖核酸高度螺旋化而呈現特定的形態,此時易為鹼性染料著色,稱之為染色體。
1970年後陸續問世的各種顯帶技術對染色體的識別作出了很大貢獻。中期染色體經過DNA變性、胰酶消化或螢光染色等處理,可出現沿縱軸排列的明暗相間的帶紋。按照染色體上特徵性的標誌可將每一個臂從內到外分為若干區,每個區又可分為若干條帶,每條帶又再分為若干個亞帶,例如“9q34.1”即表示9號染色體長臂第3區第4條帶的第1個亞帶。由於每條染色體帶紋的數目和寬度是相對恆定的,根據帶型的不同可識別每條染色體及其片段。
1980年代以來根據脫氧核糖核酸雙鏈互補的原理,套用已知序列的DNA探針進行螢光原位雜交(Fluorescenceinsituhybridization,FISH)可以識別整條染色體、染色體的1個臂、1條帶甚至一個基因,因而大大提高了染色體識別的準確性和敏感性。染色體是遺傳物質—基因的載體,控制人類形態、生理和生化等特徵的結構基因呈直線排列在染色體上。2000年6月26日人類基因組計畫(HGP)已宣布完成人類基因組序列框架圖。2001年2月12日HGP和塞雷拉公司公布了人類基因組圖譜和初步分析結果。人類基因組共有3~3.5萬個基因,而不是以往認為的10萬個。由此可見,染色體和基因二者密切相關,染色體的任何改變必然導致基因的異常。
各種生物體細胞核內染色體數目
真核生物
真核生物的基因分布住許多染色體中,一般來講這些染色體在大小上有很大不同。與細菌染色體(由環狀DNA分子構成)比較,真核染色體含有線性雙鏈DNA。DNA和多種類型的相關蛋白質構成r染色體。真核染色體的結構成分中並沒有RNA。
真核細胞中細胞核外的其他染色體(例如線粒體內的小染色體或類質粒小染色體)的數量是不固定的,可能數以千計。
進行無性生殖的物種的所有細胞中只有一套染色體,這一套染色體在所有體細胞中都是相同的。
進行有性生殖的物種的體細胞中有兩套染色體,一套來自父方;一套來自母方。而生殖細胞只有一套染色體,這一套染色體來自於具兩套染色體精原細胞或卵母細胞的減數分裂。減數分裂進行時,同大染色體(一對匹配的染色體)可能會染色體互換,由此產生與父母方的都不完全一樣,子代不是完全繼承父方或母方的新染色體。
某些生物是多倍體,體細胞有三套甚至更多套染色體。
一些植物的染色體數
| 植物物種 | # |
| 擬南芥(雙倍體) | 10 |
| 黑麥(雙倍體) | 14 |
| 玉米(雙倍體)) | 20 |
Einkornwheat(野生小麥)(雙倍體) | 14 |
| Durumwheat(四倍體) | 28 |
| 普通小麥(六倍體) | 42 |
| 馬鈴薯(四倍體) | 48 |
| 培育的菸草(雙倍體) | 48 |
| Adder'sTongueFern(雙倍體) | ~1,400 |
一些動物的染色體數(2n)
| 物種 | 數目 | 物種 | 數目 |
| 鳩鴿 | 78 | 家蠶 | 56 |
| 普通果蠅 | 8 | 幾內亞豬 | 64 |
| 蚯蚓Octodriluscomplanatus | 36 | 螺旋蝸牛 | 54 |
| 家貓 | 38 | 藏狐 | 36 |
| 實驗室的家鼠 | 40 | 家豬 | 38 |
| 兔 | 44 | 實驗室的溝鼠 | 42 |
| 野兔 | 46 | 敘利亞倉鼠 | 44 |
| 大猩猩,黑猩猩屬 | 48 | 人類 | 46 |
| 大象 | 56 | 綿羊 | 54 |
| 驢 | 62 | 牛 | 60 |
| 狗 | 78 | 馬 | 64 |
| 金魚 | 100~104 | 翠鳥 | 132 |
其它生物的染色體數目
| 物種 | 大染色體數目 | 中染色體數目 | 小染色體數目 |
| 布氏錐蟲 | 11 | 6 | ~100 |
| 雞 | 8 | 2條性染色體 | 60 |
醫學套用
生殖功能障礙者
在不孕症、多發性流產和畸胎等有生殖功能障礙的婦夫中至少有7%~10%是染色體異常的攜帶者。常見的有染色體結構異常如平衡易位和倒位以及數量異常如由於女性少一條X染色體造成的45,XO,或多一條Y染色體造成的47XXY。平衡易位和倒位由於無基因的丟失,攜帶者本身常並不發病,卻可因其生殖細胞染色體異常而導致不孕症、流產和畸胎等生殖功能障礙。性染色體數目異常除可造成不孕外,還常出現第二性徵異常。
第二性徵異常者
常見於女性,如有原發性閉經、性發育不良,伴身材矮小、肘外翻、盾狀胸和智力稍有低下,陰毛、腋毛少或缺如,後髮際低,不育等,應考慮是否有X染色體異常。常見的X染色體異常有特納氏綜合徵和環形X染色體。特納氏綜合徵患者比正常女性少一條X染色體,其染色體核型為:45,XO。環形X染色體患者由於某種原因使X染色體兩端同時出現斷裂,並在斷裂部位重接形成,環形染色體越小臨床症狀越重。早期發現這些異常並給予適當的治療可使第二性徵得到一定程度地改善,也可能獲得生育能力。
外生殖器兩性畸形者
 在成人纖維細胞上的24種(22+XY)人類染色體。
在成人纖維細胞上的24種(22+XY)人類染色體。對於外生殖器分化模糊,如陰莖伴尿道下裂,陰蒂肥大呈陰莖樣,根據生殖器外觀常難以正確決定性別的患者,通過性染色體的檢查有助於做出明確診斷。根據染色體檢查結果和臨床其它檢查,兩性畸形可分為真兩性畸形、假兩性畸形、性逆轉綜合徵等幾種不同情況。
1、真兩性畸形:內生殖器同時存在著兩性的特徵,即體內同時存在睪丸、輸精管和卵巢、輸卵管。染色體檢查表現為兩種類型:1、46,XX/46,XY,即一個機體記憶體在著兩個細胞系,每種細胞的比例決定性別取向,產生的原因:X精子和Y精子同時與兩個卵子受精後融合,或X精子和Y精子同時與卵細胞和剛形成、尚未排出卵外的極體分別受精所致。2、核型是46,XX,但是Y染色體的某些基因或片段易位於X染色體上,或常染色體基因突變而具有Y染色體的功能。
2、假兩性畸形:有進一步分為女假兩性畸形和男假兩性畸形。女假兩性畸形內生殖器表現為女性,有子宮、卵巢、輸卵管,染色體檢查為46,XX。男性假兩性畸形內生殖器表現為男性即性腺是睪丸,染色體核型是46,XY。
3、性逆轉綜合徵:即染色體核型與表型相反,例如核型是女性核型46,XX,但表型卻似男性;或核型是男性核型46,XY,但表型卻似女性。46,XX男性的主要臨床表現有睪丸發育不良,隱睪,陰莖有尿道下裂,精子少或無精子,可有喉節、鬍鬚。腋毛稀疏,群體發病率:1/2萬。46,XY女性的主要臨床表現有身材較高,卵巢為條索狀,無子宮,盲端陰道,原發性閉經,乳房不發育。
先天性多發性畸形
和智力低下的患兒及其父母染色體病的特點就是多發性畸形和智力低下,常見臨床特徵有,頭小、毛髮稀而細、眼距寬、耳位低、短頸、鼻塌而短、外生殖器發育不良、齶裂、肌張低下或亢進、顛癇、通貫掌、肛門閉鎖、身材矮小、發育遲緩、眼裂小、髮際低、持續性新生兒黃疸及明顯的青斑、眼瞼下垂、心臟畸形、腎臟畸形、虹膜或視網膜缺損等。染色體檢查可發現有21-三體綜合徵等異常。
性情異常者
身材高大、性情兇猛和有攻擊性行為的男性,有些可能為性染色體異常者。如XYY綜合徵,染色體檢查表現為比正常男性多一條Y染色體,染色體核型表現為47,XYY。患者多數表型正常,即健康情況良好,常有生育能力,但子代男性中同樣為47,XYY的機會大於正常人群。該病的發病率占一般男性人群的1/750。男性如出現身材修長、四肢細長、陰莖小、睪丸發不發育和精液中無精子者,有時還可以伴有智力異常,應通過染色體檢查確定是否患有可氏綜合症,該病患者比正常男性多一條X染色體,染色體核型表現為原`原47,XXY。其發病率在一般男性中為1/800,在男性精神發育不全者中為1%,而在男性不孕者中可高達1/10。
接觸過有害物質者
輻射、化學藥物、病毒等可以引起染色體的斷裂,如果染色體裂後原來的片段未在原來的位置上重接,將形成各種結構異常的染色體,如缺失、易位、倒位、重複、環形染色體等,這些畸變如發生在體細胞可以引起一些相應的疾病,例如腫瘤。如畸變發生在生殖細胞就發生遺傳效應,殃及子代,可以引起流產、死胎、畸形兒。
婚前檢查
婚前檢查可以發現表型正常的異常染色體攜帶者,如染色體平衡易位、倒位,染色體的平衡易位和倒位由於基因不丟失而表型正常,但極易引起流產、畸胎、死胎,盲目保胎會引起畸形兒的出生率增加。婚前檢查還可以發現表形基本正常,但性染色體異常者,這些患者可表現為性功能障礙、無生育能力等。因此,婚前檢查對優生優育有著重要的意義。
白血病及其它腫瘤患者
白血病及其它腫瘤時出現的染色體異常可使血細胞的癌基因表達,使血細胞無控制的惡性生長。不同的白血病常有各自的特徵性染色體異常,因此染色體檢查有助於白血病的診斷和預後判定。
1、慢性粒細胞白血病:Ph染色體是其標記染色體,由9號和22號染色體部份片段相互易位形成的。Ph染色體的出現為慢性粒細胞白血病的確診指標,治療過程中Ph染色體的出現或消失,還可作為療效和愈後的參考指標。
2、急性非淋巴細胞白血病:染色體改變主要為8號和21號染色體相互易位,以及15號和17號染色體相互易位,形成4條異常染色體,並且增加一條12號染色體。
3、急淋巴細胞白血病:染色體檢查可發現8號和14號染色體相互易位,4號和11號染色體相互易位,9號和22號染色體相互易位形成的6條異常染色體並增加一條21號染色體。
產生染色體異常不孕的原因
1.各種原因所致的卵母細胞或精母細胞不分離、雙精受孕、雙倍體精細胞受孕等。2.放射線、yao物、病原體、病毒等所致受精卵、胚胎染色體缺失、斷裂、環化或易位等。
3.異常染色體的遺傳及重組。
異常
原理
正常人的體細胞染色體數目為23對,並有一定的形態和結構。染色體在形態結構或數量上的異常被稱為染色體異常,由染色體異常引起的疾病為染色體病。現已發現的染色體病有100餘種,染色體病在臨床上常可造成流產、先天愚型、先天性多發性畸形、以及癌腫等。染色體異常的發生率並不少見,在一般新生兒群體中就可達0.5%~0.7%,如以我院平均每年3000新生兒出生數計算,其中可能有15~20例為染色體異常者。而在早期自然流產時,約有50%~60%是由染色體異常所致。染色體異常發生的常見原因有電離輻射、化學物品接觸、微生物感染和遺傳等。臨床上染色體檢查的目的就是為了發現染色體異常和診斷由染色體異常引起的疾病。染色體檢查是用外周血在細胞生長刺激因子——植物凝集素(PHA)作用下經37℃,72小時培養,獲得大量分裂細胞,然後加入秋水仙素使進行分裂的細胞停止於分裂中期前,以便染色體的觀察;再經低滲膨脹細胞,減少染色體間的相互纏繞和重疊,最後用甲醇和冰醋酸將細胞固定於載玻片上,在顯微鏡下觀察染色體的結構和數量;正常男性的染色體核型為44條常染色體加2條性染色體X和Y,檢查報告中常用46,XY來表示。正常女性的常染色體與男性相同,性染色體為2條XX,常用46,XX表示。46表示染色體的總數目,大於或小於46都屬於染色體的數目異常。缺失的性染色體常用O來表示。
病症示例
一、生殖功能障礙者在不孕症、多發性流產和畸胎等有生殖功能障礙的婦夫中至少有7%~10%是染色體異常的攜帶者。常見的有染色體結構變異如平衡易位和倒位以及數量異常如由於女性少一條X染色體造成的45,XO,或多一條Y染色體造成的47XXY。平衡易位和倒位由於無基因的丟失,攜帶者本身常並不發病,卻可因其生殖細胞染色體異常而導致不孕症、流產和畸胎等生殖功能障礙。性染色體數目異常除可造成不孕外,還常出現第二性徵異常。
二、第二性徵異常者
常見於女性,如有原發性閉經、性發育不良,伴身材矮小、肘外翻、盾狀胸和智力稍有低下,陰毛、腋毛少或缺如,後髮際低,不育等,應考慮是否有X染色體異常。常見的X染色體異常有特納氏綜合徵和環形X染色體。特納氏綜合徵患者比正常女性少一條X染色體,其染色體核型為:45,XO。環形X染色體患者由於某種原因使X染色體兩端同時出現斷裂,並在斷裂部位重接形成,環形染色體越小臨床症狀越重。早期發現這些異常並給予適當的治療可使第二性徵得到一定程度地改善,也可能獲得生育能力。
三、外生殖器兩性畸形者
對於外生殖器分化模糊,如陰莖伴尿道下裂,陰蒂肥大呈陰莖樣,根據生殖器外觀常難以正確決定性別的患者,通過性染色體的檢查有助於做出明確診斷。根據染色體檢查結果和臨床其它檢查,兩性畸形可分為真兩性畸形、假兩性畸形、性逆轉綜合徵等幾種不同情況。
1.真兩性畸形:內生殖器同時存在著兩性的特徵,即體內同時存在睪丸、輸精管和卵巢、輸卵管。染色體檢查表現為兩種類型:1、46,XX/46,XY,即一個機體記憶體在著兩個細胞系,每種細胞的比例決定性別取向,產生的原因:X精子和Y精子同時與兩個卵子受精後融合,或X精子和Y精子同時與卵細胞和剛形成、尚未排出卵外的極體分別受精所致。2、核型是46,XX,但是Y染色體的某些基因或片段易位於X染色體上,或常染色體基因突變而具有Y染色體的功能。
2.假兩性畸形:有進一步分為女假兩性畸形和男假兩性畸形。女假兩性畸形內生殖器表現為女性,有子宮、卵巢、輸卵管,染色體檢查為46,XX。男性假兩性畸形內生殖器表現為男性即性腺是睪丸,染色體核型是46,XY。
3.性逆轉綜合徵:即染色體核型與表型相反,例如核型是女性核型46,XX,但表型卻似男性;或核型是男性核型46,XY,但表型卻似女性。46,XX男性的主要臨床表現有睪丸發育不良,隱睪,陰莖有尿道下裂,精子少或無精子,可有喉節、鬍鬚。腋毛稀疏,群體發病率:1/2萬。46,XY女性的主要臨床表現有身材較高,卵巢為條索狀,無子宮,盲端陰道,原發性閉經,乳房不發育。
四、先天畸形和智力低下
染色體病的特點就是多發性畸形和智力低下,常見臨床特徵有,頭小、毛髮稀而細、眼距寬、耳位低、短頸、鼻塌而短、外生殖器發育不良、齶裂、肌張低下或亢進、顛癇、通貫掌、肛門閉鎖、身材矮小、發育遲緩、眼裂小、髮際低、持續性新生兒黃疸及明顯的青斑、眼瞼下垂、心臟畸形、腎臟畸形、虹膜或視網膜缺損等。染色體檢查可發現有21-三體綜合徵等異常。
五、性情異常者
身材高大、性情兇猛和有攻擊性行為的男性,有些可能為性染色體異常者。如XYY綜合徵,染色體檢查表現為比正常男性多一條Y染色體,染色體核型表現為47,XYY。患者多數表型正常,即健康情況良好,常有生育能力,但子代男性中同樣為47,XYY的機會大於正常人群。該病的發病率占一般男性人群的1/750。男性如出現身材修長、四肢細長、陰莖小、睪丸發不發育和精液中無精子者,有時還可以伴有智力異常,應通過染色體檢查確定是否患有可氏綜合症,該病患者比正常男性多一條X染色體,染色體核型表現為原`原47,XXY。其發病率在一般男性中為1/800,在男性精神發育不全者中為1%,而在男性不孕者中可高達1/10。
六、接觸過有害物質者
輻射、化學藥物、病毒等可以引起染色體的斷裂,如果染色體裂後原來的片段未在原來的位置上重接,將形成各種結構異常的染色體,如缺失、易位、倒位、重複、環形染色體等,這些畸變如發生在體細胞可以引起一些相應的疾病,例如腫瘤。如畸變發生在生殖細胞就發生遺傳效應,殃及子代,可以引起流產、死胎、畸形兒。
七、婚前檢查
婚前檢查可以發現表型正常的異常染色體攜帶者,如染色體平衡易位、倒位,染色體的平衡易位和倒位由於基因不丟失而表型正常,但極易引起流產、畸胎、死胎,盲目保胎會引起畸形兒的出生率增加。婚前檢查還可以發現表形基本正常,但性染色體異常者,這些患者可表現為性功能障礙、無生育能力等。因此,婚前檢查對優生優育有著重要的意義。
八、白血病及其它腫瘤患者
白血病及其它腫瘤時出現的染色體異常可使血細胞的癌基因表達,使血細胞無控制的惡性生長。不同的白血病常有各自的特徵性染色體異常,因此染色體檢查有助於白血病的診斷和預後判定。
1.慢性粒細胞白血病:Ph染色體是其標記染色體,由9號和22號染色體部份片段相互易位形成的。Ph染色體的出現為慢性粒細胞白血病的確診指標,治療過程中Ph染色體的出現或消失,還可作為療效和愈後的參考指標。
2.急性非淋巴細胞白血病:染色體改變主要為8號和21號染色體相互易位,以及15號和17號染色體相互易位,形成4條異常染色體,並且增加一條12號染色體。
3.急淋巴細胞白血病:染色體檢查可發現8號和14號染色體相互易位,4號和11號染色體相互易位,9號和22號染色體相互易位形成的6條異常染色體並增加一條21號染色體。
新進展
基因組研究以國際人類基因組計畫為代表,是當今生物技術研究的“熱中之熱”。人類基因組草圖的完成宣告了一個新時代——後基因組時代的到來。已經完成基因組測序的動物還有秀麗線蟲(1998年)、果蠅(2000年)、狗(2004年)和小雞(2004年)等。我國研究人員獨立完成了水稻、家蠶、雞、吸血蟲、羊等物種的全基因組測序工作。
染色體變異
名詞1、染色體變異:光學顯微鏡下可見染色體結構的變異或者染色體數目變異。
2、染色體結構的變異:指細胞內一個或幾個染色體發生片段的缺失(染色體的某一片段消失)、增添(染色體增加了某一片段)、顛倒(染色體的某一片段顛倒了180。)或易位(染色體的某一片段移接到另一條非同源染色體上)等改變。
3、染色體數目的變異:指細胞內染色體數目增添或缺失的改變。
4、染色體組:一般的,生殖細胞中形態、大小不相同的一組染色體,就叫做一個染色體組。細胞內形態相同的染色體有幾條就說明有幾個染色體組。5、二倍體:凡是體細胞中含有兩個染色體組的個體,就叫~。如.人果,蠅,玉米.絕大部分的動物和高等植物都是二倍體。
.6、多倍體:凡是體細胞中含有三個以上染色體組的個體,就叫~。如:馬鈴薯含四個染色體組叫四倍體,普通小麥含六個染色體組叫六倍體(普通小麥體細胞6n,42條染色體,一個染色體組3n,21條染色體。)。
7、一倍體:凡是體細胞中含有一個染色體組的個體,就叫~。
8、單倍體:是指體細胞含有本物種配子染色體數目的個體。
9、花葯離體培養法:具有不同優點的品種雜交,取F1的花葯用組織培養的方法進行離體培養,形成單倍體植株,用秋水仙素使單倍體染色體加倍,選取符合要求的個體作種。
套用
1、染色體變異包括染色體結構的變異(染色體上的基因的數目和排列順序發生改變),染色體數目變異。
2、多倍體育種:
a、成因:細胞有絲分裂過程中,在染色體已經複製後,由於外界條件的劇變,使細胞分裂停止,細胞內的染色體數目成倍增加。(當細胞有絲分裂進行到後期時破壞紡錘體,細胞就可以不經過末期而返回間期,從而使細胞內的染色體數目加倍。)
b、特點:營養物質的含量高;但發育延遲,結實率低。
c、人工誘導多倍體在育種上的套用:
常用方法---用秋水仙素處理萌發的種子或幼苗;
秋水仙素的作用---秋水仙素抑制紡錘體的形成;
實例:三倍體無籽西瓜(用秋水仙素處理二倍體西瓜幼苗得到四倍體西瓜;
用二倍體西瓜與四倍體西瓜雜交,得到三倍體的西瓜種子。三倍體西瓜聯會紊亂,不能產生正常的配子。)、八倍體小黑麥。
3、單倍體育種:
形成原因:由生殖細胞不經過受精作用直接發育而成。例如,蜜蜂中的雄蜂是單倍體動物;玉米的花粉粒直接發育的植株是單倍體植物。
特點:生長發育弱,高度不孕。
單倍體在育種工作上的套用常用方法:花葯離體培養法。
意義:大大縮短育種年齡。
單倍體的優點是:大大縮短育種年限,速度快,單倍體植株染色體人工加倍後,即為純合二倍體,後代不再分離,很快成為穩定的新品種,所培育的種子為絕對純種。
4、一般有幾個染色體組就叫幾倍體。如果某個體由本物種的配子不經受精直接發育而成,則不管它有多少染色體組都叫“單倍體”。
5、生物育種的方法總結如下:
①誘變育種:用物理或化學的因素處理生物,誘導基因突變,提高突變頻率,從中選擇培育出優良品種。實例---青黴素高產菌株的培育。
②雜交育種:利用生物雜交產生的基因重組,使兩個親本的優良性狀結合在一起,培育出所需要的優良品種。實例---用高桿抗鏽病的小麥和矮桿不抗鏽病的小麥雜交,培育出矮桿抗鏽病的新類型。
③單倍體育種:利用花葯離體培養獲得單倍體,再經人工誘導使染色體數目加倍,迅速獲得純合體。單倍體育種可大大縮短育種年限。
④多倍體育種:用人工方法獲得多倍體植物,再利用其變異來選育新品種的方法。(通常使用秋水仙素來處理萌發的種子或幼苗,從而獲得多倍體植物。)
實例---三倍體無籽西瓜和八倍體小黑麥的培育(6n普通小麥與2n黑麥雜交得4n後代,再經秋水仙素使染色體數目加倍至8n,這就是8倍體小黑麥)。
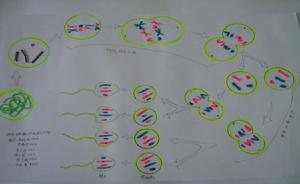
染色體配對
隨著科學技術的不斷發展,現在已經可以實現染色體配對 。配對的兩條染色體,形狀和大小一般都相同,一條來自父方,一條來自母方,叫做同源染色體。
染色體、DNA、基因的關係
①染色體與基因的關係:一條染色體上有許多基因,基因在染色體上呈直線排列。
②染色體與DNA的關係:每一條染色體上只有一個DNA分子,染色體是DNA分子的主要載體。
③DNA與基因的關係:每個DNA上有許多基因,基因是有遺傳效應的DNA片段。
細胞生活學相關知識
| 細胞生物學(cell biology)是研究細胞結構、功能及生活史的一門科學。細胞生物學由Cytology發展而來,Cytology是關於細胞結構與功能(特別是染色體)的研究。細胞生物學是以細胞為研究對象,從細胞的整體水平、亞顯微水平、分子水平等三個層次,以動態的觀點,研究細胞和細胞器的結構和功能、細胞的生活史和各種生命活動規律的學科。 |
轉基因食品相關知識
| 轉基因食品(Genetically Modified Foods,GMF)是利用現代分子生物技術,將某些生物的基因轉移到其他物種中去,改造生物的遺傳物質,使其在形狀、營養品質、消費品質等方面向人們所需要的目標轉變。以轉基因生物為直接食品或為原料加工生產的食品就是“轉基因食品”。 |
