簡介
 染色體
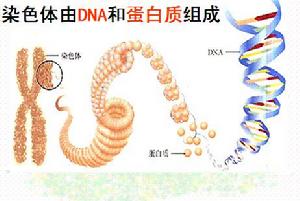
染色體 染色體是細胞核中載有遺傳信息的物質,在顯微鏡下呈圓柱狀或桿狀,主要由DNA和蛋白質組成,在細胞發生有絲分裂時期容易被鹼性染料(例如龍膽紫和醋酸洋紅)著色,因此而得名。
在無性繁殖物種中,生物體內所有細胞的染色體數目都一樣;而在有性繁殖大部分物種中,生物體的體細胞染色體成對分布,含有兩個染色體組,稱為二倍體。
性細胞如精子、卵子等是單倍體,染色體數目只是體細胞的一半。哺乳動物雄性個體細胞的性染色體對為XY,雌性則為XX。鳥類.兩棲類.爬行類和某些昆蟲的性染色體與哺乳動物不同:雄性個體的是ZZ,雌性個體為ZW。
發現
1879年德國生物學家弗萊明(Fleming·w)把細胞核中的絲狀和粒狀的物質,用染料染紅,觀察發現這些物質平時散漫地分布在細胞核中,當細胞分裂時,散漫的染色物體便濃縮,形成一定數目和一定形狀的條狀物,到分裂完成時,條狀物又疏鬆為散漫狀。
1883年美國遺傳學家、生物學家沃爾特·薩頓提出了遺傳基因在染色體上的學說。
1888年正式被命名為染色體。
1902年美國生物學家沃爾特·薩頓和鮑維里通過觀察發現細胞的減數分裂時染色體與基因具有明顯的平行關係,並推測基因位於染色體上。
1928年摩爾根通過果蠅雜交實驗證實了染色體是基因的載體,從而獲得了生理醫學諾貝爾獎。
1953年4月《自然》雜誌刊登了美國的沃森和英國的克里克在英國劍橋大學合作的研究成果:DNA雙螺旋結構的分子模型,被譽為20世紀以來生物學方面最偉大的發現。
1956年,美籍華裔遺傳學家Joe Hin Tjio(1919–2001,資料譯為莊有興或蔣有興)和Levan首次發現人的體細胞的染色體數目為46條,標誌著人類細胞遺傳學的建立。46條染色體按其大小、形態配成23對,第一對到第二十二對叫做常染色體,為男女共有,第二十三對是一對性染色體(Sex Chromosome),雄性個體細胞的性染色體對為XY;雌性則為XX。
簡介
結構
染色體的超微結構顯示染色體是由直徑僅100埃(Å,1埃=0.1納米)的DNA-組蛋白高度螺旋化的纖維所組成。每一條染色單體可看作一條雙螺旋的DNA分子。有絲分裂間期時,DNA解螺旋而形成無限伸展的細絲,此時不易為染料所著色,光鏡下呈無定形物質,稱之為染色質。有絲分裂時DNA高度螺旋化而呈現特定的形態,此時易被鹼性染料著色,稱之為常染色體。
1970年後陸續問世的各種顯帶技術對染色體的識別作出了很大貢獻。中期染色體經過DNA變性、胰酶消化或螢光染色等處理,可出現沿縱軸排列的明暗相間的帶紋。按照染色體上特徵性的標誌可將每一個臂從內到外分為若干區,每個區又可分為若干條帶,每條帶又再分為若干個亞帶,例如“9q34.1”即表示9號染色體長臂第3區第4條帶的第1個亞帶。由於每條染色體帶紋的數目和寬度是相對恆定的,根據帶型的不同可識別每條染色體及其片段。
80年代以來根據DNA雙鏈互補的原理,套用已知序列的DNA探針進行螢光原位雜交(Fluorescence in situ hybridization,FISH)可以識別整條染色體、染色體的1個臂、1條帶甚至一個基因,因而大大提高了染色體識別的準確性和敏感性。染色體是遺傳物質—基因的載體,控制人類形態、生理和生化等特徵的結構基因呈直線排列在染色體上。2000年6月26日人類基因組計畫(HGP)已宣布完成人類基因組序列框架圖。2001年2月12日HGP和塞雷拉公司公布了人類基因組圖譜和初步分析結果。人類基因組共有3~3.5萬個基因,而不是以往認為的10萬個。由此可見,染色體和基因二者密切相關,染色體的任何改變必然導致基因的異常。
成分
染色體的主要化學成份是脫氧核糖核酸(DNA)和蛋白質構成,染色體上的蛋白質有兩類:一類是低分子量的鹼性蛋白質即組蛋白(histones),另一類是酸性蛋白質,即非組蛋白蛋白質(non-histone proteins)。非組蛋白蛋白質的種類和含量不十分恆定,而組蛋白的種類和含量都很恆定,其含量大致與DNA相等。所以人們早就猜測,組蛋白在DNA·蛋白質纖絲的形成上起著重要作用。Kornberg根據生化資料,特別是根據電鏡照相,最先在1974年提出繩珠模型(beads on-a-string model),用來說明DNA·蛋白質纖絲的結構。纖絲的結構單位是核小體,它是染色體結構的最基本單位。
核小體的核心是由4種組蛋白(H2A、H2B、H3和H4)各兩個分子構成的扁球狀8聚體。我們知道,DNA分子具有典型的雙螺旋結,一個DNA分子就像是一條長長的雙螺旋的纖絲。一條染色體有一個DNA分子。DNA雙螺旋依次在每個組蛋白8聚體分子的表面盤繞約1.75圈,其長度相當於140個鹼基對。組蛋白8聚體與其表面上盤繞的DNA分子共同構成核小體。在相鄰的兩個核小體之間,有長約50~60個鹼基對的DNA連線線。在相鄰的連線線之間結合著一個第5種組蛋白(H1)的分子。密集成串的核小體形成了核質中的100埃左右的纖維,這就是染色體的“一級結構”,就像成串的珠子一樣,DNA為繩,組蛋白為珠,被稱作染色體的“繩珠模型”如圖→在這裡,DNA分子大約被壓縮了7倍。
染色體的一級結構經螺旋化形成中空的線狀體,稱為螺線體或核絲或螺線筒或螺旋管,這是染色體的“二級結構”,其外徑約300埃,內徑100埃,相鄰螺旋間距為110埃。螺旋體的每一周螺旋包括6個核小體,因此DNA的長度在這個等級上又被再壓縮了6倍。
300埃左右的螺線體(二級結構)再進一步螺旋化,形成直徑為0.4微米(μm)的筒狀體,稱為超螺旋管。這就是染色體的“三級結構”。到這裡,DNA又再被壓縮了40倍。超螺旋體進一步摺疊盤繞後,形成染色單體—染色體的“四級結構”。兩條染色單體組成一條染色體。到這裡,DNA的長度又再被壓縮了5倍。從染色體的一級結構到四級結構,DNA分子一共被壓縮了7×6×40×5=8400倍。例如,人的染色體中DNA分子伸展開來的長度平均約為幾個厘米,而染色體被壓縮到只有幾納米長。
組型
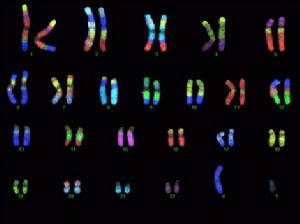
 人類的23對染色體
人類的23對染色體 染色體組型(Karyotype):描述一個生物體內所有染色體的大 小、形狀和數量信息的圖象。這種組型技術可用來尋找染色體歧變同特定疾病的關係,比如:染色體數目的異常增加、形狀發生異常變化等。以染色體的數目和形態來表示染色體組的特性,稱為染色體組型。雖然染色體組型一般是以處於體細胞有絲分裂中期的染色體的數目和形態來表示,但是,也可以其他時期,特別是以前期或分裂間期的染色體形態來表示。
關於整個染色體的情況可作下列記載而加以表示:各自的長度、粗細;著絲粒的位置;隨體及次縊痕的有無、數目、位置;凝縮部不同的部分以及異染色質部分、常染色質部分;染色粒、端粒的形態、大小及分布情況;小縊痕的數目、位置;由於溫度和藥品處理所產生的染色體分帶(band)的形態、數目、位置等等。
對於染色體組的表示,現已提出幾種方法。例如,染色體的數目是以n、2n分別表示配子和合子的染色體數目,以x表示基數,以b表示原始基數,以2x、3x、4x、……表示多倍性,以2x 1、2x-1、……等等表示非整惰性,以1、2、3、……等編號表示各個染色體。
另外,為了表示各個染色體的形態特徵,還可採用“V”形、“J”形等名稱,或者採用由A.Levan等(1964)所提出的根據著絲粒的位置進行分類的方法等。關於人類的染色體組型的表示法,在國際上是統一的(在丹佛1960、倫敦1963、芝加哥1966、巴黎1971等地召開的人類染色體會議上所制訂的),已規定了為了表示染色體形態特徵的染色體臂比、著絲點指數等指標。
DNA序列
染色體要確保在細胞世代中保持穩定,必須具有自主複製、保證複製的完整性、遺傳物質能夠平均分配的能力,與這些能力相關的結構序列是:
自主複製
20世紀70年代末首次在酵母菌中發現。自主複製DNA序列具有一個複製起始點,能確保染色體在細胞周期中能夠自我複製,從而保證染色體在世代傳遞中具有穩定性和連續性。
著絲粒
著絲粒DNA序列與染色體的分離有關。著絲粒DNA序列能確保染色體在細胞分裂時能被平均分配到2個子細胞中去。
著絲粒DNA序列特點:
(1)一方面在所有的真核生物中它們的功能是高度保守的,另一方面即使在親緣關係非常相近的物種之間它們的序列也是多樣的。
(2)絕大多數生物的著絲粒都是由高度重複的串聯序列構成的,然而,在著絲粒的核心區域,重複序列的刪除,擴增以及突變發生的非常頻繁,種種研究表明,重複序列並不是著絲粒活性所必須的。
(3)有些科學家提出了可能是DNA的二級結構甚至是高級結構是決定著絲粒位置和功能的因素。即功能的序列無關性。
端粒
為一段短的正向重複序列,在人類為TTAGGG的高度重複序列。端粒DNA功能是保證染色體的獨立性和遺傳穩定性。
![染色體[細胞核的組成部分]](/img/6/511/nBnauM3X4cTN0gDN0EDM4UjM2UTM1QDN5MjM5ADMwAjMwUzLxAzL2YzLt92YucmbvRWdo5Cd0FmLyE2LvoDc0RHa.jpg) 染色體[細胞核的組成部分]
染色體[細胞核的組成部分] 染色體的分裂分三種; 一是母鍾分裂,這個一般發生在受精卵的早期,人類具體就是從一條受精卵分裂為個體的23對染色體的過程,意思是按照母體藍圖進行子代分裂,被分裂的23對染色體分別可以造出各種組織器官,如果第一條是造肝的,那么它上面的所有造肝的基因片段都被打開,相反其它器官的製造信息都被關閉,這個過程母體藍圖染色體要分裂4次(按幾何級數分裂);
二是子鍾分裂,按照母體藍圖分裂的23對人類染色體已經在“母鍾分裂”過程中分別被打開,它們各自按照各自的“子代藍圖”進行下面造器官的分裂,一個個有機的器官從此被造出,並且開始發揮各自的功能,這個過程子體藍圖染色體要分裂24次(個物種染色體的不同,其分裂的次數也不同,不過一個總的原則是按染色體數分裂),在24次分裂後,一個完整的人體就被造出來;三是孫鍾分裂,一個獨立的人體,在生長發育的過程中,還有一些器質性和功能性的東西沒有出現,所以必須再打開,進行再分裂。比如七歲兒童脫牙,十多歲少年具有生育能力,有些遺傳病到一定時候的發作,等等。
對應三種分裂,必須有三種控制分裂發生的手段。母鍾分裂是“端點(又叫端粒)控制體系”,這種分裂的原始觸發點在外界,比如飄蕩在空氣中的細菌,它只要沒有接觸食物或易感物,就永遠是不產生分裂的原命(見百度詞條“雙命”),一旦接觸,在端點的作用下就開始母鍾分裂。子鍾分裂是受制於子鍾染色體的端點,與外界刺激無關。孫鍾染色體分裂受制於染色體外相對應的一些蛋白質,它們的功能僅僅是到一定時間將這個包含某信息的片段打開。
依此看來,染色體就是人體的生物鐘。所以我們將第一條受精卵叫“母鍾”,將母鍾分裂出來的23對染色體叫子鍾,將23對染色體造出的各種組織器官所包含的染色體叫“孫鍾”,改變子鍾、孫鐘的染色體都不可以改變遺傳,只有改變母鐘的基因才可以造成“變異”。
染色體可以攜帶“遺傳基因”但是不能傳遞“打開信息”,打開某個基因段的所有信息都是通過染色體端點或染色體外的蛋白質發揮作用才完成分裂或複製的。分裂是染色體整體的,複製是染色體某個基因片段的。
不同狀態
基因和染色體
基因在細胞里並非一盤“散沙”或“散兵游勇”,它們大多有規律地集中在細胞核內的染色體上,而且每一種生物細胞內染色體的形態和數目都是一定的。
染色體複製時
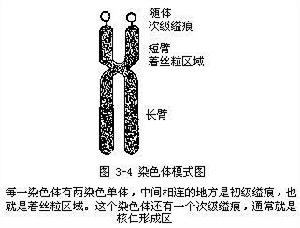 染色體模式圖
染色體模式圖 染色體在複製以後,含有縱向並列的兩個染色單體(c hroma-tids),只有在著絲粒(centromere)區域仍聯在一起。著絲粒在染色體上的位置是固定的。由於著絲粒位置的不同,把染色體分成大致相等或長短不等的兩臂(arms)。著絲粒的位置在染色體中間或中間附近時,染色體兩臂的長度差不多,這著絲粒叫做中間著絲粒或亞中間著絲粒。著絲粒的位置靠近染色體的一端時,根據著絲粒離開端部的遠近,這著絲粒叫做近端部著絲粒或端部著絲粒。著絲粒所在的地方往往表現為一個縊痕,所以著絲粒又稱初級縊痕(primary constriction)。
有些染色體上除了初級縊痕以外,還有一個次級縊痕(secon-dary constriction),連上一個叫做隨體(satellite)的遠端染色體小段。次級縊痕的位置也是固定的。在細胞分裂將結束時,核內出現一個到幾個核仁,核仁總是出現在次級縊痕的地方,所以次級縊痕也叫做核仁形成區(nucleolar organizer)如圖:著絲粒是有絲分裂或減數分裂中的染色體高度壓縮的一個區域,在此紡錘體纖維與其結合。複雜的DNA序列構成了著絲粒。發育的麵包酵母(啤酒酵母)的著絲粒長度約為220個鹼基對,並且通過多種與剩餘DNA結合組蛋白有明顯區別的蛋白質共同保護其免受限制性核酸內切酶(簡稱限制酶)的消化。雖然著絲粒區域通過特殊的蛋自質保護其免受限制性內切核酸酶的攻擊,但該區域投有核小體而且被去凝聚,這似乎說明了在有絲分裂和減數分裂過程中著絲粒區域被高度縮窄原因。著絲粒的220對鹼基序列兩側是限制性內切核酸酶敏感位點,該位點的功能也許是促進DNA的斷裂,有助於染色單體在後期的相互分離。限制性內切核酸酶是一種在核酸內特殊位點進行切割的酶類。
不同生物
真核生物
真核生物的基因分布在許多染色體中,一般來講這些染色體在大小上有很大不同。與細菌染色體(由環狀DNA分子構成)比較,真核染色體含有線性雙鏈DNA。DNA和多種類型的相關蛋白質構成r染色體。真核染色體的結構成分中並沒有RNA。
真核染色體可被不同程度的濃縮。最低的濃縮結構是伸展的核小體形式(extended nucleosome form)(圖3一la)。核小體是由近乎球狀的組蛋白形成的八聚體(histone)(H2a、H2b、H3和H4)和在其外圍繞兩圈的DNA所構成。DNA的約200個鹼基對圍繞著由組蛋白構成的球形體。並有DNA的50個鹼基對連線相鄰的核小體。雖然真核染色體伸展的核小體形式與細菌當中看到的串珠樣的結構相似,但是這些結構可能並不相同。伸展的核小體形式存在於染色體將被複製的區域,或存在於與RNA分子合成有關的區域。真核DNA更緊密的狀態是螺線管形式(solenoid form)。與核小體結合的組蛋白H1誘導其組裝成6個核小體的環,並且這些環組成圓筒狀螺線管結構。在分裂間期大多數真核染色體以螺線管形式存在。進行複製或被表達的(轉錄為RNA分子)這些部分去解凝成為伸展的核小體形式。DNA的複製發生在間期的S階段。基因表達發生在間期的所有階段(G0、S和G1)。那些不進行增殖的(進入細胞周期)真核細胞被認為是在G0階段,並且與間期細胞相似。在間期細胞核中看到的染色質是由絕大多數以螺線管形式存在的DNA 構成。
DNA最緊密的狀態是環狀的螺線管形式。DNA結合蛋白促進螺線管在支架蛋白中心核前後形成環狀。在一些真核生物中螺線管的l8個環組成了一個盤狀結構。染色體凝聚為數百個疊在一起的盤狀結構。在有絲分裂和減數分裂的過程中,可觀察到環狀的螺線管形式。由於許多長的染色體必須在細胞內移動,並且在移動過程中可能被牽扯,所以染色體的濃縮是必要的。
原核細菌
在高中階段,染色體必須由DNA和蛋白質組成。由於原核生物的染色體一般不含組蛋白,所以高中階段稱原核生物沒有染色體。
一般而言,原核生物的染色體可以進行複製,但大多數細胞容易存活多份。
所有必需的細菌基因存在於細胞質中的單個環狀雙鏈DNA(dsDNA)染色體中。細菌染色體與質膜相附著。細菌染色體(bacterial chromosome)依其種類不同可編碼1000個或5000 個蛋白質。除了細菌染色體以外,還可有一個或多個較小的染色體,稱為質粒,它一般指定 20一100個蛋白質。質粒是環狀雙鏈DNA分子,它可與或不與質膜附著。質粒編碼的大多數或全部蛋白質在正常環境條件下並不是細胞生存所絕對必需的。許多質粒編碼的蛋白質使其把一些遺傳信息向其他細胞轉移成為可能,並促進稀有化合物的代謝。或使細胞可抵抗某些化學物質或重金屬。
由DNA、蛋白質和RNA構成的細菌染色體是高度濃縮的。它不僅通過拓撲異構酶(topoisomerase)形成超螺旋,並且,環繞在由RNA和蛋白質形成的“核”的內外。許多DNA的負電荷被多胺[如精胺(spermine)和亞精胺(spermidine)]和DNA纏繞著的鹼性蛋白質所中和。通過柔和地裂解細菌細胞得到的DNA外觀呈串珠狀。雖然細菌染色體也是高度濃縮的,但是,在光學顯微鏡下它們不能被看到。在透射電子顯微鏡下,細菌染色體的外觀與非分裂的真核細胞核內的染色質(chiromatin)非常相像。
人類
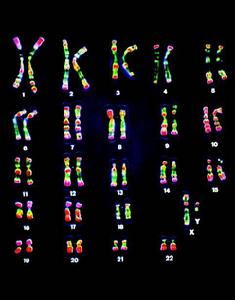
 人類的染色體
人類的染色體 男性體細胞染色體的 組成:22對常染色體+XY。女性體細胞染色體的組成:22對常染色體+XX。但是,男性生殖細胞染色體的組成:22條常染色體+X或Y。女性生殖細胞染色體的組成:22條常染色體+X。
人體的體細胞內有23對染色體。包括22對常染色體和一對性染色體。性染色體包括:X染色體和Y染色體。含有一對X染色體的受精卵發育成女性,而具有一條X染色體和一條Y染色體者則發育成男性。這樣,對於女性來說,正常的性染色體組成是XX,男性是XY。這就意味著,女性細胞減數分裂產生的配子都含有一個X染色體;男性產生的精子中有一半含有X染色體,而另一半含有Y染色體。精子和卵子的染色體上攜帶著遺傳基因,上面記錄著父母傳給子女的遺傳信息。同樣,當染色體異常時,就可形成遺傳性疾病。男性不育症中因染色體異常引起者約占2%~21%,尤其以少精子症和無精子症多見。
數目列表
每一種生物的染色體數是恆定的。多數高等動植物是二倍體(diploid),也就是說,每一身體細胞中有兩組同樣的染色體(有時與性別直接有關的染色體,即性染色體,可以不成對)。親本的每一配子帶有一組染色體,叫做單倍體(haploid),用n來表示。兩個配子結合後,具有兩組染色體,叫做二倍體,用2n表示。例如玉米的二倍體染色體數是20(2n=20),即有10對染色體,如圖→。人的染色體數是46(2n=46),即有23對染色體,如圖←。但多數微生物的營養體是單倍體,例如鏈孢霉的單倍體染色體數是7。
| 通名 | 學名 | 雙倍體數 |
| 動物 | ||
| 人類 | Homo sapiens | 46 |
| 彌猴 | Macacamalatta | 42 |
| 黃牛 | Bostaurus | 60 |
| 豬 | Susscrofa | 38 |
| 狗 | Canis familiaris | 78 |
| 貓 | Felis domesticus | 38 |
| 馬 | Equus Calibus | 64 |
| 驢 | Equus asinus | 62 |
| 山羊 | Capara hircus | 60 |
| 綿羊 | Ovis aries | 54 |
| 小家鼠 | Mus musculus | 40 |
| 大家鼠 | Rattus norvegicus | 42 |
| 水貂 | Mustela vison | 30 |
| 豚鼠 | Cavia cobaya | 64 |
| 兔 | Oryctolagus cuniculus | 44 |
| 家鴿 | pigeon | 約80 |
| 雞 | Gallus domesticus | 約78 |
| 火雞 | Meleagris gallopavo | 約80 |
| 鴨 | Anas platyrhyncho | 約80 |
| 家蠶 | Bombyx mori | 56 |
| 家蠅 | Musca domestica | 12 |
| 果蠅 | Drosophila melanogaster | 8 |
| 蜜蜂 | Apis mellifera | ♀32♂16 |
| 蚊 | Culex pipiens | 6 |
| 佛蝗 | Phlaeobainfumata | ♀24♂23 |
| 水螅 | hydra | 32 |
| 植物 | ||
| 洋蔥 | Alliumcepa | 16 |
| 大麥 | Hordeum uulgare | 14 |
| 水稻 | Oryza sativa | 24 |
| 小麥 | Triticum vulgare | 42 |
| 玉米 | Zea mays | 20 |
| 金魚草 | Antirrhinum majus | 16 |
| 陸地棉 | Gossypium hirsutum | 52 |
| 中棉 | Gossypium arboreum | 26 |
| 豌豆 | Pisum sativum | 14 |
| 香豌豆 | Lathyrus odoratus | 14 |
| 蠶豆 | Vicia faba | 12 |
| 菜豆 | phaseolus vulgaris | 22 |
| 向日葵 | Helianthusannuus | 34 |
| 菸草 | Nicotiana taldcum | 48 |
| 番茄 | Solanum lycopersicum | 24 |
| 松 | Pinus species | 24 |
| 青菜 | Brassica chinensis | 20 |
| 甘藍 | Brassica oleracea | 18 |
| 月見草 | Oenothera biennis | 14 |
| 微生物 | ||
| 鏈孢霉 | Neurospora crassa | 7 |
| 青黴菌 | Penicillium species | 4 |
| 麴黴 | Aspergillus nidulans | 8 |
| 衣藻 | Chlamydomonas reinhardi | 16 |
性染色體
 染色體
染色體 遺傳的染色體學說的證據來自於這樣的實驗,一些特殊基 因的遺傳行為和性染色體(Sex Chromosome)傳遞的關係。性染色體在高等真核生物的兩種性別中是不同的。性染色體的發現為Sutton-Boveri的學說提供了一個實驗證據。
在孟德爾以前(1891年)德國的細胞學家亨金(Henking,H)曾經用半翅目的昆蟲蝽做實驗,發現減數分裂中雄體細胞中含11對染色體和一條不配對的單條染色體,在第一次減數分裂時,它移向一極,亨金無以為名,就稱其為“X”染色體。後來在其它物種的雄體中也發現了“X”染色體。
1900年麥克朗(McClung, C.E)等就發現了決定性別的染色體。他們採用的材料多為蚱蜢和其它直翅目昆蟲。1902年麥克朗發現了一種特殊的染色體,稱為副染色體(accessory chromosome)。在受精時,它決定昆蟲的性別。1906年威爾遜(Wilson, E.B)觀察到另一種半翅目昆蟲(Proteror)的雌體有6對染色體,而雄性只有5對,另外加一條不配對的染色體,威爾遜稱其為X染色體,其實雌性是有一對性染色體,雄性為XO型。
在1905年斯蒂文斯(Stevens, N)發現擬步行蟲屬(Tenebrio molitor)中的一種甲蟲雌雄個體的染色體數目是相同的,但在雄性中有一對是異源的,大小不同,其中有一條雌性中也有,但是是成對的;另一條雌性中怎么也找不到,斯蒂文斯就稱之為Y染色體。在黑腹果蠅中也發現了相同的情況,果蠅共有4對染色體,在雄性中有一對是異形的染色體。在1914年塞勒(Seiler,J)證明了在雄蛾中染色體都是同形的,而在雌蛾中有一對異形染色體。他們根據異形染色體的存在和性別的相關性,發現了性染色體,已完全證實了他們的推論是完全正確的。嚴格地說異形染色體的存在僅是一條線索,而不是證據,不能因為存在異形染色體,就表明其為性染色體。一定要通過實驗證明這條染色體上存在決定性別的主要基因,方能定論。
X-染色體
2005年3月17日,在Nature雜誌上發表的一篇文章宣告基本完成對人類X染色體的全面分析。對X染色體的詳細測序是英國Wellcome Trust Sanger研究中心領導下世界各地多所著名學院超過250位基因組研究人員共同完成的,是人類基因組計畫的一部分。
從屬於NIH的美國國家人類基因組研究院的負責人弗朗西絲·柯林斯博士(Francis S. Collins, Ph.D)表示“對X染色體的詳細研究成果代表了生物學和醫藥學領域進展的一個新的里程碑。新的研究確認了X染色體上有1098個蛋白質編碼基因--有趣的是,這1098個基因中只有54個在對應的Y染色體上有相應功能。
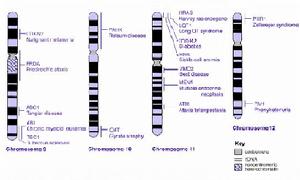
染色體研究是臨床遺傳學研究的基礎。測序結果表明X染色體包涵多達1100種基因。但另人吃驚的是,與之相關的疾病也有百餘種,如X染色體易碎症、血友病、孤獨症、肥胖肌肉萎縮病和白血病等。看來這條染色體決不容小視!
X染色體對應的另一半就是Y染色體。人類Y染色體的測序工作也已經完成,並且發現它並沒有人們之前想像的那樣脆弱。Y染色體上有一個“睪丸”決定基因則對性別決定至關重要。已經知道的與Y染色體有關的疾病有十幾種。
異常
原理
正常人的體細胞染色體數目為23對,並有一定的形態和結構。染色體在形態結構或數量上的異常被稱為染色體異常,由染色體異常引起的疾病為染色體病。現已發現的染色體病有100餘種,染色體病在臨床上常可造成流產、先天愚型、先天性多發性畸形、以及癌腫等。染色體異常的發生率並不少見,在一般新生兒群體中就可達0.5%~0.7%,如以我院平均每年3000新生兒出生數計算,其中可能有15~20例為染色體異常者。而在早期自然流產時,約有50%~60%是由染色體異常所致。染色體異常發生的常見原因有電離輻射、化學物品接觸、微生物感染和遺傳等。臨床上染色體檢查的目的就是為了發現染色體異常和診斷由染色體異常引起的疾病。
染色體檢查是用外周血在細胞生長刺激因子——植物凝集素(PHA)作用下經37℃,72小時培養,獲得大量分裂細胞,然後加入秋水仙素使進行分裂的細胞停止於分裂中期前,以便染色體的觀察;再經低滲膨脹細胞,減少染色體間的相互纏繞和重疊,最後用甲醇和冰醋酸將細胞固定於載玻片上,在顯微鏡下觀察染色體的結構和數量;正常男性的染色體核型為44條常染色體加2條性染色體X和Y,檢查報告中常用46,XY來表示。正常女性的常染色體與男性相同,性染色體為2條XX,常用46,XX表示。46表示染色體的總數目,大於或小於46都屬於染色體的數目異常。缺失的性染色體常用O來表示。
病症示例
1. 生殖功能障礙者
在不孕症、多發性流產和畸胎等有生殖功能障礙的夫婦中至少有7%~10%是染色體異常的攜帶者。常見的有染色體結構變異如平衡易位和倒位以及數量異常如由於女性少一條X染色體造成的45,XO,或多一條Y染色體造成的47XXY。平衡易位和倒位由於無基因的丟失,攜帶者本身常並不發病,卻可因其
染色體異常而導致不孕症、流產和畸胎等生殖功能障礙。性染色體數目異常除可造成不孕外,還常出現第二性徵異常。
2. 第二性徵異常者
常見於女性,如有原發性閉經、性發育不良,伴身材矮小、肘外翻、盾狀胸和智力稍有低下,陰毛、腋毛少或缺如,後髮際低,不育等,應考慮是否有X染色體異常。常見的X染色體異常有特納氏綜合徵和環形X染色體。特納氏綜合徵患者比正常女性少一條X染色體,其染色體核型為:45,XO。環形X染色體患者由於某種原因使X染色體兩端同時出現斷裂,並在斷裂部位重接形成,環形染色體越小臨床症狀越重。早期發現這些異常並給予適當的治療可使第二性徵得到一定程度地改善,也可能獲得生育能力。
3. 外生殖器兩性畸形者
對於外生殖器分化模糊,如陰莖伴尿道下裂,陰蒂肥大呈陰莖樣,根據生殖器外觀常難以正確決定性別的患者,通過性染色體的檢查有助於做出明確診斷。根據染色體檢查結果和臨床其它檢查,兩性畸形可分為真兩性畸形、假兩性畸形、性逆轉綜合徵等幾種不同情況。
(1)真兩性畸形:內生殖器同時存在著兩性的特徵,即體內同時存在睪丸、輸精管和卵巢、輸卵管。染色體檢查表現為兩種類型:1、46,XX/46,XY,即一個機體記憶體在著兩個細胞系,每種細胞的比例決定性別取向,產生的原因:X精子和Y精子同時與兩個卵子受精後融合,或X精子和Y精子同時與卵細胞和剛形成、尚未排出卵外的極體分別受精所致。2、核型是46,XX,但是Y染色體的某些基因或片段易位於X染色體上,或常染色體基因突變而具有Y染色體的功能。
(2)假兩性畸形:有進一步分為女假兩性畸形和男假兩性畸形。女假兩性畸形內生殖器表現為女性,有子宮、卵巢、輸卵管,染色體檢查為46,XX。男性假兩性畸形內生殖器表現為男性即性腺是睪丸,染色體核型是46,XY。
(3)性逆轉綜合徵:即染色體核型與表型相反,例如核型是女性核型46,XX,但表型卻似男性;或核型是男性核型46,XY,但表型卻似女性。46,XX男性的主要臨床表現有睪丸發育不良,隱睪,陰莖有尿道下裂,精子少或無精子,可有喉節、鬍鬚。腋毛稀疏,群體發病率:1/2萬。46,XY女性的主要臨床表現有身材較高,卵巢為條索狀,無子宮,盲端陰道,原發性閉經,乳房不發育。
4.先天畸形和智力低下
染色體病的特點就是多發性畸形和智力低下,常見臨床特徵有,頭小、毛髮稀而細、眼距寬、耳位低、短頸、鼻塌而短、外生殖器發育不良、齶裂、肌張低下或亢進、顛癇、通貫掌、肛門閉鎖、身材矮小、發育遲緩、眼裂小、髮際低、持續性新生兒黃疸及明顯的青斑、眼瞼下垂、心臟畸形、腎臟畸形、虹膜或視網膜缺損等。染色體檢查可發現有21-三體綜合徵等異常。
5. 性情異常者
身材高大、性情兇猛和有攻擊性行為的男性,有些可能為性染色體異常者。如XYY綜合徵,染色體檢查表現為比正常男性多一條Y染色體,染色體核型表現為47,XYY。患者多數表型正常,即健康情況良好,常有生育能力,但子代男性中同樣為47,XYY的機會大於正常人群。該病的發病率占一般男性人群的1/750。男性如出現身材修長、四肢細長、陰莖小、睪丸發不發育和精液中無精子者,有時還可以伴有智力異常,應通過染色體檢查確定是否患有克氏綜合症,該病患者比正常男性多一條X染色體,染色體核型表現為原`原47,XXY。其發病率在一般男性中為1/800,在男性精神發育不全者中為1%,而在男性不孕者中可高達1/10。
6. 接觸過有害物質者
輻射、化學藥物、病毒等可以引起染色體的斷裂,如果染色體裂後原來的片段未在原來的位置上重接,將形成各種結構異常的染色體,如缺失、易位、倒位、重複、環形染色體等,這些畸變如發生在體細胞可以引起一些相應的疾病,例如腫瘤。如畸變發生在生殖細胞就發生遺傳效應,殃及子代,可以引起流產、死胎、畸形兒。
7. 婚前檢查
婚前檢查可以發現表型正常的異常染色體攜帶者,如染色體平衡易位、倒位,染色體的平衡易位和倒位由於基因不丟失而表型正常,但極易引起流產、畸胎、死胎,盲目保胎會引起畸形兒的出生率增加。婚前檢查還可以發現表形基本正常,但性染色體異常者,這些患者可表現為性功能障礙、無生育能力等。因此,婚前檢查對優生優育有著重要的意義。
8. 白血病及其它腫瘤患者
白血病及其它腫瘤時出現的染色體異常可使血細胞的癌基因表達,使血細胞無控制的惡性生長。不同的白血病常有各自的特徵性染色體異常,因此染色體檢查有助於白血病的診斷和預後判定。
(1)慢性粒細胞白血病:Ph染色體是其標記染色體,由9號和22號染色體部份片段相互易位形成的。Ph染色體的出現為慢性粒細胞白血病的確診指標,治療過程中Ph染色體的出現或消失,還可作為療效和愈後的參考指標。
(2)急性非淋巴細胞白血病:染色體改變主要為8號和21號染色體相互易位,以及15號和17號染色體相互易位,形成4條異常染色體,並且增加一條12號染色體。
(3)急淋巴細胞白血病:染色體檢查可發現8號和14號染色體相互易位,4號和11號染色體相互易位,9號和22號染色體相互易位形成的6條異常染色體並增加一條21號染色體。
新進展
基因組研究以國際人類基因組計畫為代表,是當今生物技術研究的“熱中之熱”。人類基因組草圖的完成宣告了一個新時代——後基因組時代的到來。已經完成基因組測序的動物還有秀麗線蟲(1998年)、果蠅(2000年)、狗(2004年)和小雞(2004年)等。
我國研究人員獨立完成了水稻、家蠶、雞、吸血蟲、羊等物種的全基因組測序工作。
染色體變異
名詞
1、染色體變異:光學顯微鏡下可見染色體結構的變異或者染色體數目變異。
2、染色體結構的變異:指細胞內一個或幾個染色體發生片段的缺失(染色體的某一片段消失)、增添(染色體增加了某一片段)、顛倒(染色體的某一片段顛倒了180)或易位(染色體的某一片段移接到另一條非同源染色體上)等改變。
3、染色體數目的變異:指細胞內染色體數目增添或缺失的改變。
4、染色體組:一般的,生殖細胞中形態、大小不相同的一組染色體,就叫做一個染色體組。細胞內形態相同的染色體有幾組就說明有幾個染色體組。
5、二倍體:凡是體細胞中含有兩個染色體組的個體,就叫~。如.人,果蠅,玉米.絕大部分的動物和高等植物都是二倍體。
6、多倍體:凡是體細胞中含有三個以上染色體組的個體,就叫~。如:馬鈴薯含四個染色體組叫四倍體,普通小麥含六個染色體組叫六倍體(普通小麥體細胞6n,42條染色體,一個染色體組3n,21條染色體。)。
7、一倍體:凡是體細胞中含有一個染色體組的個體,就叫一倍體。
8、單倍體:是指體細胞含有本物種配子染色體數目的個體。
9、花葯離體培養法:具有不同優點的品種雜交,取F1的花葯用組織培養的方法進行離體培養,形成單倍體植株,用秋水仙素使單倍體染色體加倍,選取符合要求的個體作種。
套用
1、染色體變異包括染色體結構的變異(染色體上的基因的數目和排列順序發生改變),染色體數目變異。
2、多倍體育種:
a、 成因:細胞有絲分裂過程中,在染色體已經複製後,由於外界條件的劇變,使細胞分裂停止,細胞內的染色體數目成倍增加。(當細胞有絲分裂進行到後期時破壞紡錘體,細胞就可以不經過末期而返回間期,從而使細胞內的染色體數目加倍。)
b、 特點:營養物質的含量高;但發育延遲,結實率低。
c、人工誘導多倍體在育種上的 套用:
常用方法---用秋水仙素處理萌發的種子或幼苗;
秋水仙素的作用---秋水仙素抑制紡錘體的形成;
實例:三倍體無籽西瓜(用秋水仙素處理二倍體西瓜幼苗得到四倍體西瓜;
用二倍體西瓜與四倍體西瓜雜交,得到三倍體的西瓜種子。三倍體西瓜聯會紊亂,不能產生正常的配子。)、八倍體小黑麥。
3、單倍體育種:
形成原因:由生殖細胞不經過受精作用直接發育而成。例如,蜜蜂中的雄蜂是單倍體動物;玉米的花粉粒直接發育的植株是單倍體植物。
特點:生長發育弱,高度不孕。
單倍體在育種工作上的 套用常用方法:花葯離體培養法。
意義:大大縮短育種年齡。
單倍體的 優點是:大大縮短育種年限,速度快,單倍體植株染色體人工加倍後,即為純合二倍體,後代不再分離,很快成為穩定的新品種,所培育的種子為絕對純種。
4、一般有幾個染色體組就叫幾倍體。如果某個體由本物種的配子不經受精直接發育而成,則不管它有多少染色體組都叫“單倍體”。
5、生物育種的方法總結如下:
①誘變育種:用物理或化學的因素處理生物,誘導基因突變,提高突變頻率,從中選擇培育出優良品種。實例---青黴素高產菌株的培育。
②雜交育種:利用生物雜交產生的基因重組,使兩個親本的優良性狀結合在一起,培育出所需要的優良品種。實例---用高桿抗鏽病的小麥和矮桿不抗鏽病的小麥雜交,培育出矮桿抗鏽病的新類型。
③單倍體育種:利用花葯離體培養獲得單倍體,再經人工誘導使染色體數目加倍,迅速獲得純合體。單倍體育種可大大縮短育種年限。
④多倍體育種:用人工方法獲得多倍體植物,再利用其變異來選育新品種的方法。(通常使用秋水仙素來處理萌發的種子或幼苗,從而獲得多倍體植物。)
實例---三倍體無籽西瓜和八倍體小黑麥的培育(6n普通小麥與2n黑麥雜交得4n後代,再經秋水仙素使染色體數目加倍至8n,這就是8倍體小黑麥)。
染色體的起源
染色體起源是細胞核起源的核心過程,但依然還是未解之謎。迄今為止的學說主要有:共營模型(syntrophic model)、自演化模型(autogenous model)、病毒性真核生物起源模型(viral eukaryogenesis model)、外膜假說(exomembrane hypothesis)、壓縮和結構化假說(packing and structurization hypothesis),等等。
共營模型認為,與現代產甲烷古菌類似的某些古老的古菌,侵入並生活在類似於現代粘細菌的細菌體內,形成了早期的細胞核。古菌與真核生物在特定蛋白質(如組蛋白)基因的相似性被認為是支持以古菌為基礎的細胞核起源理論的證據。
自演化模型認為原真核(proto-eukaryotic)細胞直接自細菌演化而來,並不需要通過內共生。證據來自一類專性好氧菌——浮黴菌(Planctomycete),它們具有清晰的胞內膜結構,其中,有一種稱之為Gemmata obscuriglobus的出芽菌,其染色質被雙層的核膜所包裹,類似於真核生物的核的結構,而斯氏小梨形菌(Pirellula staleyi)的核被單層的細胞質內膜ICM所包裹。但是,這一模型並未進一步解釋核實如何形成的。
病毒性真核生物起源模型認為,病毒感染原核生物導致了膜結合的細胞核與其他真核生物特徵的產生。證據是真核生物和病毒在大分子結構上存在一定相似性,譬如,線性DNA鏈、mRNA的加帽,以及與蛋白質的緊密結合(病毒的外套膜類似於組蛋白)。該假說的其中一種觀點認為,吞噬作用形成了早期的細胞“捕食者”,並隨之演化出細胞核。
外膜假說認為,細胞核是起源自演化出第二層外細胞膜的單個早期細胞,而包裹原來細胞的內膜則轉變成了核膜,並逐漸演化出精巧的核孔結構,以便於將內部(如核糖體亞基)合成的物質送出核外。
壓縮與結構化假說認為,細胞核源自原核細胞基因組的大型化(包括DNA的複製錯誤或多倍化、側向基因轉移方式、內共生融合等)。核的成型及有絲分裂的出現主要是為了滿足將巨大的DNA分子準確地分配到子代中去的需求,因此,如何將長鏈DNA有效地壓縮(藉助組蛋白)成若干染色體以及如何將多個染色體同時分離(藉助紡錘體)是核演化的關鍵。從原核生物到真核生物,基因組的DNA總量大約增加了3.5個數量級,這與現代真核生物的DNA壓縮比(packing ratio)驚人地一致。包括核膜在內的細胞內膜系統就是為了實現對複雜生化系統進行秩序化管控,或者說,秩序化是通過細胞內部的模組化得以實現的。
