特徵
 粘菌門
粘菌門粘菌門是介於動植物之間的一類生物,約有500種。它們的生活史中一般是動物性的,另一段是植物性的。
營養體是一團裸露的原生質體,多核,無葉綠體,能作變形蟲式運動,與動物相似。生殖時能產生具纖維素壁的孢子,為植物性狀。
粘菌在生長期或營養期為裸露的無細胞壁多核的原生質團,稱變形體(plasmodium),其營養體構造、運動或攝食方式與原生動物中的變形蟲相似,但在繁殖時期產生具纖維質細胞壁的孢子,又具真菌的性狀。
事實上粘菌是介於動物和真菌之間的生物。大多數生於森林中陰暗和潮濕的地方,在腐木上、落葉上或其他濕潤的有機物上。大多數粘菌為腐生菌,無直接的經濟意義,只有極少數粘菌寄生在經濟植物上,危害寄主。
繁殖
變形體粘菌生活於陰濕環境中的朽木敗葉中。它們是一團裸露、多核的原生質體,一般直徑約幾個厘米,能呈變形蟲狀爬行。變形體能吞食細菌、酵母菌、真菌孢子、有機物碎塊,並在原生質內消化吸收。在良好條件下,粘菌變形體
 粘菌門
粘菌門不斷增長,細胞核也不斷分裂,整個變形體可重達幾百克,覆蓋面積可達1平方米。從變形體結構、運動、營養方式看,它與原生動物中的變形蟲相似,是動物。
當粘菌生活條件不良時,變形體就分成若干小團,每個小團形成若干個直立的突起,然後每個突起都發育成一個有細柄的孢子囊,進入生殖時期。這時,孢子囊里的每個細胞核進行減數分裂,原生質也分割成許多塊原生質體,縮成球形,分泌纖維素的壁,變成孢子。粘菌的孢子成熟後,由乾裂的孢子囊中散出,在乾燥環境裡可生存很久。當環境適宜時,每個孢子萌發成1或4個有鞭毛的單倍性遊動細胞。遊動細胞兩兩配合,成為一個二倍性合子,合子不經休眠,立即發育成一個多核的變形體。從粘菌的生殖期來看,有孢子囊,孢子有纖維素構成的壁,這些又是植物的特徵。
生活史
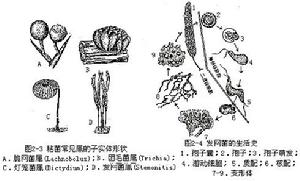
粘菌的營養階段為自由生、無細胞壁、多核的變形蟲狀原生質所組成的原質團。處於營養階段的原質團,具負趨光性,在黑暗潮濕的環境中,以細菌、酵母菌和其他有機顆粒為食。營養耗盡時,向有光處遷移,並且就在光暗交界處向子實體階段過渡,原質團中所含的無數核(二倍體的)進行同步有絲分裂,形成孢子囊和孢子。孢子成熟後,從囊中釋出,在潮濕的表面上萌發,生出遊動孢子。遊動孢子可兩兩結合,成為二倍體合子。許多合子集聚在一起,又形成多核原質團。
主要類群
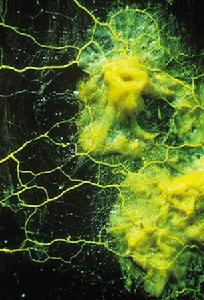
粘菌在全世界約有500種,一般分為3個綱,即粘菌綱(Myxomycetes)、集胞[粘]菌綱(Acrasiomycetes)
 粘菌門
粘菌門(一)粘菌綱
有真正的變形體,通常產生具鞭毛的遊動細胞,子實體的外表有1層包被包圍著孢子。
本綱最常見的為髮網菌屬(Stemonitis),其營養體為裸露的原生質團,稱變形體。變形體呈不規則的網狀,直徑數厘米,在陰濕處的腐木上或枯葉上緩緩爬行。在繁殖時,變形體爬到乾燥光亮的地方,形成很多的髮狀突起,每個突起發育成1個具柄的孢子囊(子實體)。孢子囊通常長筒形,紫灰色,外有包被(peridium)。孢子囊柄伸入囊內的部分,稱囊軸(columella),囊內有孢絲(capilltium)交織成孢網。然後原生質團中的許多核進行減數分裂,原生質團割裂成許多塊單核的小原生質,每塊小原生質分泌出細胞壁,形成1個孢子,藏在孢絲的網眼中。成熟時,包被破裂,藉助孢網的彈力把孢子彈出。
孢子在適合的環境下,即可萌發為具2條不等長鞭毛的遊動細胞。遊動細胞的鞭毛可以收縮,使遊動細胞變成1個變形體狀細胞,稱變形菌胞。由遊動細胞或變形菌胞兩兩配合,形成合子,合子不經過休眠,
 粘菌門
粘菌門(二)根腫菌綱
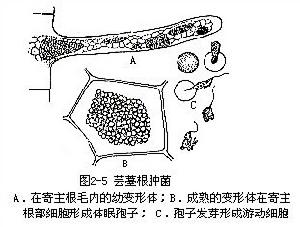
本綱菌類是寄生於高等植物、藻類或真菌上的粘菌。在整個生活史中,大部分生活在寄主細胞內,其營養組織為原生質團,不形成子實體;其休眠孢子單個或成團、無壁或在某些種內包以薄壁,這些休眠孢子直接在寄主細胞內形成。
芸薹根腫菌(PlasmodiophorabrassicaeWoronin)可作為本綱的代表。該菌侵害十字花科植物根部使患根腫病。芸薹根腫菌的生活大部分在寄主根部細胞中渡過,寄主死後,在病部細胞中形成休眠孢子。孢子微小,單核,單孢子放出後,在適當的條件下,即可萌發為遊動細胞,從十字花科植物的根毛侵入,不久失去鞭毛變為變形菌胞,變形菌胞的核重複分裂,形成1個多核的原生質團,在寄主根部細胞中,最後形成休眠孢子。
代表菌類
髮網菌
 髮網菌
髮網菌髮網菌(Stemonitis)是粘菌中最常見的種類,其變形體呈不規則網狀,直徑數厘米,能藉助體形的改變在陰濕處的腐木或枯葉上緩慢爬行,並能吞食固體食物。在繁殖時,變形體爬到乾燥光亮的地方,形成很多髮狀突起,每個突起發育成一個具柄的孢子囊(子實體)。孢子囊通常長筒形,外有包被(peridium)。孢子囊柄伸入囊內的部分,稱囊軸(columella),囊內有孢絲(capillitium)交織成孢網。然後原生質團中的許多核進行減數分裂,原生質團割裂成許多塊單核的小原生質,每塊小原生質分泌出細胞壁,形成1個孢子,藏在孢絲的網眼中。成熟時,包被破裂,藉助孢網的彈力把孢子彈出。
分布
粘菌的分布是世界性的,在有植物或植物殘體而且溫度適宜的地方都可存在。溫帶種類最多,熱帶或高寒山區很少,南極有記載。
在生物界的地位
粘菌的起源和親緣關係,迄今仍不明確。從它的特性來看是屬於動物和真菌之間的;就其
 燈籠菌
燈籠菌1949年,粘菌學家馬丁(G.W·Martin)認為,粘菌是從1種與原生動物相類似的祖先進化而來的,在他的“真菌分區中”,列為粘菌綱。1950年,貝西(E.A.Bessey)認為粘菌是動物,稱菌形動物(Mycetozoa),並正式把粘菌分到動物界的原生動物門內。
因為學者對粘菌的地位、起源和分類的看法很不一致,通常採用施羅特(Schroter)1889年的傳統分類法,把粘菌和細菌、真菌分開,成為植物界中的1個獨立門,稱粘菌門(Myxothal-lophyta)。安茲沃斯等於1973年將粘菌門(Myxomycota)隸屬於真菌界。
粘菌的套用
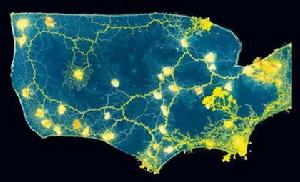 粘菌生長形成的美國交通網路圖
粘菌生長形成的美國交通網路圖粘菌在尋找食物時生長延伸著卷鬚,在食物來源之間收縮外部卷鬚尋找最有效的路徑。英國研究人員基於計算機模型,通過培育黏菌成功地複製了美國當地道路和鐵路的路徑。 在模型中的城市部分放置著壓縮食物,而在鄉鎮部分放置著較少的營養物質,這樣黏液菌會在模型中的城市部分生長得更加旺盛,為了吸收營養物質黏菌會在荒地之間形成清晰的“交通路線”。這些黏液菌能夠均勻地遍布美國地圖模型上,並適應性地沿著地形結構蔓延生長。由於黏液菌的培育生長具有很強的“彈性”,它還可以用於創建移動通訊和交通運輸網路的模型。
