![爬蟲[爬行類動物]](/img/6/fad/nBnauM3X3YTOwcjMycDN3AjNyMTM3AjMygTM2IzMwAzMwIzL3QzL1AzLt92YucmbvRWdo5Cd0FmLxE2LvoDc0RHa.jpg "爬蟲[爬行類動物]")
定義
 鱷魚圖片
鱷魚圖片傳統的林奈分類系統里,爬行動物的層級為綱,名稱是爬行綱(Reptilia)。但由於該分類沒有包含恐龍及似哺乳爬行動物的現存後代——鳥類及哺乳類,而使其成為並系群。根據親緣分支分類法,現代爬行動物與鳥類組合成的單系群分類是蜥形綱,再與合弓綱組成單系群羊膜動物。
現存的爬行動物包含四個目:
鱷目(Crocodilia):包含鱷魚、長吻鱷、短吻鱷、以及凱門鱷等23個種。
喙頭蜥目(Sphenodontia):包含生存於紐西蘭的喙頭蜥,共2個種。
有鱗目(Squamata):包含蜥蜴、蛇、以及蚓蜥,接近7900個種。
龜鱉目(Testudines):包含海龜與陸龜,接近300個種。
現代的爬行動物棲息於每個大陸,除了南極洲以外,但它們主要分布於熱帶與副熱帶地區。現存的爬行動物,體型最大的是鹹水鱷,可達7米以上,最小的是侏儒壁虎(Jaraguasphaero),只有1.6厘米長。除了少數的龜鱉目以外,所有的爬行動物都覆蓋者鱗片。
雖然所有的細胞在代謝時都會產生熱量,但大部分的爬行動物不能產生足夠的熱量以保持體溫,因此被稱為冷血動物或變溫動物(棱皮龜可能是個例外)。爬行動物依靠環境來吸收或散發內部的熱量。例如,在向陽處或陰暗處之間移動,或藉由循環系統將溫暖血液流動至身體內部,將較冷血液流動至身體表層。大部份生存於天然棲息地的爬行動物,可將身體內部的體溫維持在相當狹窄的變化範圍內。不同於兩棲類,爬行動物的表皮厚,因此不需要棲息在水邊,吸取水分。由於體溫調節方面的關係,爬行動物可以較少的食物維生。溫血動物通常以較快速度移動,某些蜥蜴、蛇、或鱷魚的移動速度較快。
大部分的爬行動物是卵生動物,它們的胚胎由羊膜所包覆。然而,某些有鱗目能夠直接生下後代,例如以卵胎生或胎生的方式。大部分的胎生爬行動物藉由不同形式的胎盤給予胎兒養分,類似哺乳類的方式。它們通常提供給剛出生幼體適當的初步親代養育。
爬蟲的分類
分類歷史
在1758年,瑞典生物學家卡爾·林奈(CarlvonLinné)建立雙名法後幾年,他根據部分蛇、蜥蜴生存於水中,而建立兩棲綱(Amphibia),以包含兩棲動物、爬行動物。在1768年,自然學家Josephus Nicolaus Laurenti建立爬行綱(Reptilia),但當時兩棲動物、爬行動物的界定仍然模糊;Repere在拉丁語中意為“爬行”。直到18世紀末期,兩棲動物、爬行動物才被視為不同的動物。在1825年,拉特雷耶·皮埃爾·安德烈(PierreAndréLatreille)建立“Batracia”綱,以取代兩棲綱,並將四足動物分成四大類:兩棲動物、爬行動物、鳥類、哺乳動物,已經相當接近現代傳統分類法。
在托馬斯·亨利·赫胥黎(ThomasHenryHuxley)的推廣下,安德烈的分類逐漸普及。理查·歐文(RichardOwen)則將似哺乳爬行動物包含在爬行綱的範圍。
在傳統的分類法中,爬行動物的範圍為羊膜動物除去鳥類與哺乳類。因此爬行綱(Reptilia)被定義為包含鱷魚、短吻鱷、喙頭蜥、蜥蜴、蛇、蚓蜥、以及烏龜在內的動物群。這個名稱與定義成為後來常見的用法。然而在最近幾年,許多分類學家主張生物學分類應該為單系群,這些分類應該包含特定物種的所有後代。上述的爬行綱定義因為不包含哺乳綱與鳥綱,而被視為並系群,它們都是原始爬行動物的後代。
哺乳類是個演化支,因此親緣分支分類法承認傳統的哺乳綱分類;而鳥類也是個演化支,正式的名稱為鳥綱。事實上,哺乳綱與鳥綱是羊膜動物的次演化支。但傳統分類法中的爬行綱並不是一個演化支,它其實是部分的羊膜動物,不包含哺乳綱與鳥綱。爬行綱並沒有可以鑑定的共有衍征。爬行綱的定義方式包含擁有與缺乏的特徵:缺乏羽毛與毛髮的羊膜動物。換句話說,親緣分支分類法學家提出,傳統的爬行綱是“非鳥類、也並非哺乳類的羊膜動物”。
在1916年,Edwin Stephen Goodrich建立了蜥形綱(Sauropsida)與獸形綱(Theropsida),用來區別蜥蜴、鳥類與其近親,以及哺乳類與其已滅絕近親。兩個演化支的心臟與血管差異,以及前腦結構等特徵,證實這個分類法。根據Goodrich宣稱,蜥形綱與獸形綱共同從一個較早期的原始動物演化而來,名為“Protosauria”,該分類單元也包含某些古生代的兩棲類以及早期爬行動物。
在1956年,David Meredith Seares Watson發現這兩個分類單元在爬行動物歷史的非常早期就開始分開演化,因此他將“Protosauria”分為蜥形綱與獸形綱。他也分別將鳥類與哺乳類排除在蜥形綱與獸形綱以外。因此根據他的說法,蜥形綱包含:前棱蜥目、始鱷目、米勒古蜥科、龜鱉目、有鱗目、喙頭目、鱷目、槽齒目、恐龍、翼龍目、魚龍目、以及鰭龍超目。
Watson的分類法補充了Goodrich版本的盲點,但從未廣為接受。根據美國古生物學家阿爾弗雷德·羅默(AlfredSherwoodRomer)的著作Vertebrate Paleontology,依照頭顱骨的顳顬孔數量與不同位置,爬行綱分為四個亞綱,分別為:
無孔亞綱(Anapsida):沒有顳顬孔。
單孔亞綱(Synapsida):一個下方顳顬孔。目前被稱為合弓綱。
調孔亞綱(Euryapsida):一個上方顳顬孔。目前被包含於雙孔亞綱之內。
雙孔亞綱(Diapsida):兩個顳顬孔。
以上四個亞綱,除了單孔亞綱以外,目前都屬於蜥形綱。
分類學
以下目級列表是根據班頓在2004年的研究:
羊膜動物Amniota
合弓綱Synapsida
盤龍目Pelycosauria
獸孔目Therapsida
哺乳綱Mammalia
蜥形綱Sauropsida
無孔亞綱Anapsida
龜鱉目Testudines
雙孔亞綱Diapsida
纖肢龍目Araeoscelidia
楊氏蜥形目Younginiformes
魚龍超目Ichthyosauria
鱗龍形下綱Lepidosauromorpha
鰭龍超目Sauropterygia
楯齒龍目Placodontia
幻龍目Nothosauroidea
蛇頸龍目Plesiosauria
鱗龍超目Lepidosauria
喙頭目Sphenodontida
有鱗目Squamata
主龍形下綱Archosauromorpha
原蜥形目Prolacertiformes
主龍類Archosauria
鑲嵌踝類主龍Crurotarsi
植龍目Phytosauria
堅蜥目Aetosauria
勞氏鱷目Rauisuchia
鱷形超目Crocodylomorpha
鱷目Crocodylia
Avemetatarsalia
鳥頸類主龍Ornithodira
翼龍目Pterosauria
恐龍總目Dinosauria
鳥臀目Ornithischia
蜥臀目Saurischia
鳥綱Aves
系統發生學
以下簡易版的爬行動物演化樹:
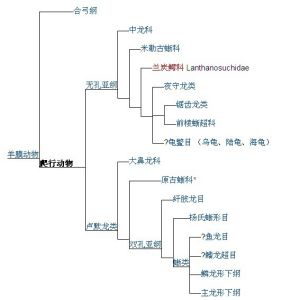 演化樹
演化樹演化歷史
最早期的爬行動物,出現於石炭紀晚期,約3億2000萬3億1000萬年前,演化自迷齒亞綱的爬行形類(Reptiliomorpha)。林蜥是已知最古老的爬行動物,身長約20到30厘米,化石發現於加拿大的新斯科細亞省。西洛仙蜥(Westlothiana)曾被認為是最早的爬行動物,但目前被認為較接近於兩棲類,而非羊膜動物。油頁岩蜥與中龍都為最早期的爬行動物之一。最早期的爬行動物生存於石炭紀晚期的沼澤森林,但體型小於同時期的迷齒螈類,例如原水蠍螈(Proterogynrius)、Cochleosaurus。石炭紀末期的小型冰河期,使得早期爬行動物有機會成長至較大的體型。
演化支
在最早的爬行動物出現後不久,出現了兩個演化支,一個是無孔亞綱。無孔亞綱擁有堅硬的頭顱骨,沒有顳顬孔,僅有與鼻孔、眼睛、脊椎相對應的洞孔。烏龜被認為是目前僅存的無孔動物,因為它們擁有相同的頭顱骨特徵;但最近有些科學家認為烏龜是返祖遺傳到原始的狀態,以增加它們的保護能力。關於烏龜的起源,目前有副爬行動物的後代、失去顳顬孔的雙孔亞綱兩派學說。
另一群演化支是雙孔亞綱,頭顱骨上有兩個顳顬孔,位於眼睛後方。雙孔動物進一步分化為兩個支系:鱗龍類包含現代蜥蜴、蛇、喙頭蜥,可能還有中生代的已滅絕海生爬行動物;主龍類包含現代鱷魚與鳥類,以及已滅絕的翼龍目與恐龍。
而最早期、具堅硬頭顱骨的羊膜動物也演化出另一獨立的演化支,稱為單孔亞綱。合弓動物的眼睛後方有一對窩孔,可減輕頭顱骨重量,並提供頜部肌肉附著點,增加咬合力。單孔亞綱最後演化為哺乳類,因此被稱為似哺乳爬行動物。單孔亞綱過去為爬行綱的一個亞綱,但目前為個別的合弓綱。
二疊紀
在石炭紀末期,合弓類、爬行類動物成為陸地優勢動物。迷齒螈類、爬行形類仍然生存在水邊,而合弓類盤龍目首先演化至較大的體型,例如基龍與異齒龍。在二疊紀中期,氣候多次變遷,造成生態系統的改變,獸孔目取代盤龍目,成為陸地優勢動物。
無孔類爬行動物繁盛於二疊紀。其中,鋸齒龍類也演化出較大的體型。大部分無孔類爬行動物在二疊紀—三疊紀滅絕事件滅絕,龜鱉類可能是它們的後代。
在二疊紀時期,雙孔類爬行動物並不繁盛、體型小。但在二疊紀末期演化出兩個演化支:主龍形下綱、鱗龍形下綱,最後演化出大部分的現存爬行動物。
中生代
二疊紀末期的二疊紀—三疊紀滅絕事件,造成合弓類動物、無孔類爬行動物的大量滅絕,而主龍形下綱成為陸地優勢動物。早期主龍類已具有直立的四足步態,在短期內演化出多種演化支:恐龍、翼龍目、鱷形超目、以及其他三疊紀的主龍類。其中,恐龍是三疊紀後期到白堊紀末期的陸地優勢動物群。因此中生代有時被戲稱為“恐龍時代”、“爬行動物時代”。在侏羅紀中期,獸腳亞目恐龍演化出許多有羽毛恐龍,更進一步演化出鳥類。
相對於主龍形下綱,鱗龍形下綱則可能演化出多群海生爬行動物:楯齒龍目、幻龍目、蛇頸龍目、滄龍科;魚龍類可能演化自更原始的雙孔類爬行動物。鱗龍形下綱也演化出多種陸棲小型爬行動物,例如喙頭蜥、蜥蜴、蛇、蚓蜥。
在恐龍的競爭壓力下,獸孔目演化出體型小、高代謝率的物種,並在侏羅紀晚期演化出哺乳動物。
新生代
白堊紀末期的白堊紀—第三紀滅絕事件,使恐龍、翼龍目、大部分海生爬行動物、大部分鱷形類滅絕,而鳥類、哺乳動物在新生代再次繁盛、多樣化,因此新生代被戲稱為“哺乳動物時代”。只有龜鱉類、喙頭蜥、蜥蜴、蛇、蚓蜥、鱷魚繼續存活到現代,主要生存於熱帶與副熱帶地區。現存爬行動物大約有8200個種,其中半數屬於蛇。
生物學
循環系統
大部分的現存爬行動物具有閉合的循環系統,它們具有三腔室心臟,由兩個心房與一個心室所構成,心室的分割方式並不一致。它們通常只有一對大主動脈。當它們的血液流經三腔室心臟時,含氧血與缺氧血只有少量混合。但是,血液可改變流通方式,缺氧血可流向身體,含氧血可流向肺臟,使爬行動物的體溫調節更有效率,尤其是水生物種。
鱷魚具有四腔室心臟,可以在水中以三腔室心臟運作。某些蛇類與蜥蜴(例如巨蜥與蟒蛇),具有三腔室心臟,但可以四腔室心臟方式運作。因為它們心臟的皮瓣可在心動周期時,隨者擴張、收縮隔開心室。某些喙頭蜥可藉由皮瓣,產生類似哺乳動物與鳥類的心臟運動。
呼吸系統
所有的爬行動物都用肺臟呼吸。水生烏龜發展出具滲透性的皮膚,某些爬行動物可用泄殖腔來增加氣體交換的面積。即使具有這些構造,它們仍需要肺臟來完成呼吸作用。在各個主要的爬行動物種類之間,完成肺部換氣的方式並不相同。有鱗目爬行動物的肺部幾乎只藉由軸肌換氣,而這個肌肉也同時用於運動。因為這個限制,大部分的有鱗目爬行動物必須在激烈的移動中停止呼吸。但是,有些爬行動物找到了解決方法。巨蜥以及一些其他的蜥蜴物種使用一種使用到喉嚨肌肉的方法來補足原本的“軸呼吸”。這使得這些動物可以在激烈運動中為肺部完全充氣,所以可以持續進行有氧呼吸很長一段時間。據了解,雙領蜥具有原始橫隔膜可將胸腔和腹腔分開來。即使此膜不能動,它還是能借著將內臟的重量和肺分開而增加肺的吸氣量。鱷魚其實擁有和哺乳類動物相似的有肌肉橫隔膜。不同的是,鱷魚的橫隔膜將恥骨拉回(部分的骨盆,在鱷魚中能移動),從而使肝下降,以騰出空間使肺能脹大。這種橫隔膜式的機制被稱作“肝活塞”。
關於海龜和陸龜如何呼吸已經被研究了很多次。迄今為止,只有少數物種已經被深入研究到足以了解其如何呼吸。其結果表明,海龜及陸龜分別找到了不同的方法來解決這個問題。但問題是,大部分的龜殼是僵硬的,且不能像其他羊膜動物使用的膨脹和收縮來為肺部換氣。一些烏龜,如印度鱉(Lissemyspunctata)有一層肌肉包覆著肺部。當它收縮時,烏龜可以呼氣;在靜止時,烏龜可以收回四肢進到體腔內,並強制使空氣排出肺部。而當烏龜伸出其四肢時肺部內的壓力會減少,烏龜因此可以吸進空氣。烏龜的肺連線到殼內的上方,而肺的底部附著在其餘內臟(由結締組織連線)。藉由一系列特殊的肌肉(大約相當於一個橫隔膜),因為許多這些肌肉的附著點都在他們的前肢上(事實上,很多的肌肉在收縮時擴張到四肢的袋子裡),烏龜能夠推動它們的內臟向上或向下,造成有效的呼吸。現今已研究了三個物種在運動期間的呼吸,並且顯示了它們之間不同的模式。當成年雌性綠蠵龜沿著要產卵的海灘爬行時並不呼吸。它們在陸上運動時屏住呼吸,只有在停頓休息時呼吸。北美的箱龜運動期間還是繼續呼吸,而且其換氣的周期並不與肢體動作相關。它們可能是利用其腹部肌肉在運動中呼吸。最後被研究的物種是紅耳龜。它們也在運動中呼吸,但它們在運動中的呼吸比在運動間停頓時的呼吸還少,顯示肢體運動和呼吸器具之間可能有機械干擾。箱龜也被觀察到在完全密封的殼內呼吸。
骨骼系統
爬行動物的骨骼系統大多數由硬骨組成,骨骼的骨化程度高,很少保留軟骨部分。
大部分的爬行動物缺乏次生顎,所以當它們進食時,無法同時呼吸。鱷魚已發展出骨質次生顎,使它們可在半隱沒至水中時持續呼吸,並防止嘴中的獵物掙扎時,傷及腦部。石龍子科也演化出骨質次生顎。
排泄系統
爬行動物的排泄系統主要藉由兩顆腎臟組成。雙孔動物所排泄的主要含氮廢物是尿酸;而烏龜主要排泄尿素,類似哺乳類。不像哺乳類與鳥類的腎臟,爬行動物的腎臟不能夠製造尿液,尿液可以儲藏更多的身體廢物。這是因為它們缺乏鳥類與哺乳類所擁有的腎元,稱為亨利環(loopofHenle)。因此許多爬行動物使用它們的結腸與排泄腔來再度吸收水分。有些爬行動物甚至可利用膀胱來取得水分。有些爬行動物的鼻部與舌部具有鹽腺,可以儲存額外的鹽分。
神經系統
爬行動物與兩棲類的腦部具有相同的基本部分,但它們的大腦比小腦稍大。爬行動物的感覺器官多發展良好,除了少部份物種,例如蛇缺乏外耳,但仍具有中耳與內耳。爬行動物具有12對腦神經。
繁衍
除了陸龜與海龜以外,大部分的雄性爬行動物具有成對的管狀性器官,稱為半陰莖。陸龜與海龜則具有單一陰莖。所有的龜鱉目皆為卵生動物,而某些蜥蜴與蛇是卵胎生或胎生動物。爬行動物藉由泄殖腔來交配、繁衍;泄殖腔位於尾巴基部,可用來排泄與繁殖。
對於某些有鱗目物種而言,雌性個體若達到一定的數量,它們便能自行複製出單性染色體。這種無性繁殖方式稱為孤雌生殖(parthenogenesis),目前已有六科蜥蜴,以及一種蛇,被確認具有這種無性繁殖能力,例如:數種壁虎、美洲蜥蜴科(尤其是Aspidocelis)以及蜥蜴科。被封閉的科莫多龍(巨蜥科)也具有孤雌生殖能力。
除了以上物種以外,變色龍、飛蜥科、夜蜥蜴科、以及嵴尾蜥科,也被推測具有孤雌生殖能力。
爬行動物的蛋,外部是鈣質蛋殼或皮革,覆蓋者內部的羊膜、羊膜囊、以及尿囊。
