 平衡恐爪龍的想像圖
平衡恐爪龍的想像圖馳龍科(Dromaeosauridae)又譯奔龍科,是群類似鳥類的獸腳亞目恐龍。它們是群小型、細長的肉食性恐龍,繁盛於白堊紀。在非正式的用法裡,它們通常被稱為“盜龍”,這個名稱因為電影《侏羅紀公園》(Jurassic Park)出現的伶盜龍而開始盛行。馳龍科的屬名在希臘文意為“奔跑的蜥蜴”,dromeus(δρομευς) 意為“奔跑”,而sauros(σαυρος) 意為“蜥蜴”。
馳龍科的化石被發現於北美洲、歐洲、北非、日本、中國、蒙古、馬達加斯加、阿根廷、以及南極洲等地。它們首次出現於中侏羅紀巴通階,約1億6700萬年前,並且存活到白堊紀末的馬斯垂克階,約6550萬年前,存活時間超過1億年。侏羅紀的馳龍科化石僅發現牙齒,但沒有發現過身體部份的化石。
科學分類
界: 動物界Animalia
門: 脊索動物門 Chordata
綱: 蜥臀目 Sauropsida
總目: 恐龍總目 Dinosauria
目: 蜥臀目 Saurischia
亞目: 獸腳亞目 Theropoda
下目: 恐爪龍下目 Deinonychosauria
科: 馳龍科 Dromaeosauridae
敘述
 伶盜龍的骨架,位於布魯塞爾
伶盜龍的骨架,位於布魯塞爾身體架構
馳龍科的獨特身體架構,有助於恐龍是種活躍且快速動物的理論,以及恐龍與鳥類是近親的理論。在羅伯特·巴克(Robert T. Bakker)替約翰·奧斯特倫姆(John Ostrom)的1969年專題論文所做的圖解中將恐爪龍描繪成快速奔跑的姿態,這成為史上最具影響力的古生物學重建圖。馳龍科具有大型頭部、鋸齒邊緣牙齒、狹窄口鼻部、眼睛向前,顯示具有一定程度的立體視覺。如同大部分的獸腳類恐龍,馳龍科的頸部長,呈S狀彎曲,身體相當短。它們的手臂長,某些物種的手臂可以在胸前交叉,手部相當大,上有三根長手指,第一指最短,第二指最長,手指上有大型指爪。恥骨末端大,往尾巴基部延伸。第二腳趾上有大型、彎曲趾爪。尾巴修長,尾椎長而低平,第14節尾椎之後缺乏橫突(Transverse process)與神經棘。
目前已發現部分馳龍科的身體覆蓋者羽毛,包含翼與尾巴上的大型、片狀羽毛,可能所有的馳龍科都有羽毛。這個理論是在80年代中晚期首次出現,並在1999年得到證實。這些改變導致馳龍科在電影與重製模型中的形象不斷改變。
鑑定特徵
馳龍科具有以下特徵:額骨短而呈T字形,構成上顳孔的前緣、方骨後突接觸方軛骨、背椎的椎體橫突對(Parapophyses)豎起、獨特的第二腳趾、人字形骨與尾椎的前關節突延長,延伸至另外幾節脊椎、喙骨上有關節盂窩。
體型
馳龍科是群小型到中型的恐龍,身長範圍從0.5公尺的小盜龍,到超過6公尺的猶他盜龍與阿基里斯龍。目前所發現最原始的馳龍科是大黑天神龍,身長只有70公分。這個新發現顯示近鳥類的共同祖先可能非常的小,身長約65公分,體重約600到700公克;近鳥類包含馳龍科、傷龍科、以及鳥類。
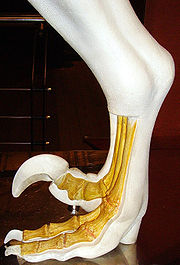 馳龍科的腳部構造模型
馳龍科的腳部構造模型腳部
如同其他獸腳類恐龍,馳龍科以後肢行走。然而,它們只有用第三、第四腳趾支撐身體重量,從足跡化石顯示,它們的而第二趾明顯地後縮。後縮的第二腳趾上有大型、鐮刀狀、彎曲趾爪,被認為是作為殺死獵物使用。
尾巴
馳龍科具有長尾巴。大部分的尾椎有骨質、棒狀前關節突,某些物種的尾巴則具有骨化肌腱。在奧斯特倫姆的恐爪龍研究中,他提出這些特徵將尾巴硬挺,不易彎曲,惟有尾巴基部可靈活活動。但是,蒙古伶盜龍的一個保存良好標本(編號IGM 100/986),尾巴向後彎曲呈S狀,且呈天然狀態。這顯示它們的尾巴可做出一定程度的平行運動。馳龍科的尾巴被假設在奔跑或滑翔時,作為穩定器或平衡器使用;在顧氏小盜龍與中國鳥龍的一個標本(編號 NGMC 91)的尾巴末端,有羽毛構成的小型、鑽石形扇狀物,可能作為氣動性上的穩定器,以及方向舵使用。
與鳥類的關係
馳龍科屬於獸腳亞目,與早期的鳥類有許多共同特徵。有許多研究討論過馳龍科與鳥類的關係,隨者許多新證據的出現,馳龍科與鳥類關係的假設也不斷改變。在2001年,馬克·諾瑞爾(Mark Norell)與其同事研究大量的虛骨龍類化石,並提出馳龍科是鳥類的最近親,兩者與傷齒龍科共同屬於初鳥類。諾瑞爾更指出,馳龍科是主鳥類中的一個並系群。在2002年,S.H. Hwang與其同事參考了諾瑞爾的研究與新發現化石,提出鳥類是馳龍科、傷齒龍科兩者的近親。
古生物學界目前的共識多同意Hwang等人的研究,馳龍科與傷齒龍科是最近親,兩者共同組成恐爪龍下目。而恐爪龍下目是鳥類的姊妹分類單元,也是鳥類的最近親。但古生物學界還沒有足夠證據可指出馳龍科(以及它們的祖先)能否飛行或滑翔。
相關理論
馳龍科傳統上被認為是非鳥類的恐龍,然而有些研究人員認為馳龍科也屬於鳥綱演化支。首先,馳龍科具有羽毛,符合傳統的鳥類定義-具有羽毛。羽龍意為“隱藏的飛行者”,它們最初因其具有羽毛而被命名為鳥類,造成許多誤解。
目前至少兩派研究人員提出馳龍科是從可飛行的祖先演化而來。其中一派主張鳥類先演化出現,喬治·奧利舍夫斯基(George Olshevsky)被認為是這理論的首位提出者。葛瑞格利·保羅(Gregory S. Paul)則提出許多證據,指出馳龍科演化自會飛行的恐龍,可能類似始祖鳥。根據此理論,體型較大的馳龍科是在演化過程中失去飛行能力,如同紐西蘭的已滅絕恐鳥、以及駝鳥。在1988年,保羅提出馳龍科與現代鳥類的關係,比始祖鳥與現代鳥類的關係更為接近。但在2002年,保羅改提出馳龍科是始祖鳥的最近親。
在2002年,S.H. Hwang與其同事提出小盜龍是最原始的馳龍科。在2003年,徐星與其同事根據小盜龍的羽毛、翅膀等特徵,提出馳龍科的祖先能夠滑翔。而體型較大的馳龍科則在演化過程中失去滑翔能力,成為陸棲動物。
在2002年,斯特芬·柯瑞克斯(Stephen Czerkas)敘述、命名羽龍。柯瑞克斯錯誤地判斷羽龍的羽毛只位在前肢與手部,將羽龍重建成雙翼恐龍;並認為羽龍較接近鳥類,而非滑翔動物。但是,日後有其他科學家提出羽龍可能是小盜龍的次異名。而柯瑞克斯也在他的著作中更正他的錯誤。
另一派研究人員,例如賴利·馬丁(Larry Martin),則提出馳龍科與所有手盜龍類都不是恐龍。馬丁並提出鳥類與手盜龍類並非近親,但在2004年,馬丁修正他的理論,認為手盜龍類是鳥類的近親。但馬丁認為手盜龍類在演化過程中失去飛行能力,而鳥類演化自非恐龍的主龍類,而獸腳亞目的大部分成員並不屬於恐龍。
 瑟馬普利斯始祖鳥標本,具有類似馳龍科的第二趾爪。
瑟馬普利斯始祖鳥標本,具有類似馳龍科的第二趾爪。在2005年,Mayr與Peters敘述了一個在懷俄明州瑟莫普利斯鎮發現的良好始祖鳥標本,發現始祖鳥的生理結構比原先認知的更類似恐龍。另外,他們還具有類似馳龍科的大幅彎曲第二趾爪、原始上顎、未反轉後趾。Mayr與Peters的種系發生學研究提出孔子鳥與小盜龍的親緣關係更為接近,而離始祖鳥較遠,使初鳥綱成為並系群演化支。他們更提出其近鳥類祖先能夠飛行或滑翔,而馳龍科與傷齒龍科隨後失去飛行或滑翔能力。Corfe與Butler則以研究方法層面提出反對意見。
在2007年,另有其他的理論出現。艾倫·特納(Alan Turner)與其同事命名了一種新馳龍科-大黑天神龍,大黑天神龍是最基礎、原始的馳龍科,比小盜龍更原始。大黑天神龍具有短手臂,無法滑翔。特納等人認為飛行能力演化自初鳥類,而馳龍科的祖先無法飛行、滑翔。根據親緣分支分類法研究,原始的大黑天神龍顯示馳龍科的祖先無法飛行。
分類系統
 恐爪龍(大)與鷲龍(小)的骨骼,芝加哥菲爾德博物館
恐爪龍(大)與鷲龍(小)的骨骼,芝加哥菲爾德博物館分類學
在1992年,W.D. Matthew與巴納姆·布郎(Barnum Brown)建立了馳龍亞科,當時只包含馳龍屬,屬於現在已廢止的恐爪龍科。馳龍科與傷齒龍科共同組成恐爪龍下目。
馳龍科的各亞科經常因為新的研究而改變成員內容,但大致上包含以下各生物群。而且許多成員的保存狀態不好,無法分類到任一亞科之中,例如最基底的大黑天神龍。
半鳥亞科是馳龍科中最基礎的亞科。半鳥亞科是個充滿謎題的生物群,是最少得到理論支持的亞科,其中有些或全部的成員可能並不屬於馳龍科。其中較大、生活於地面的成員,例如鷲龍與半鳥,顯示出強烈的飛行適應,但它們可能過大而不能飛行。另一個成員脅空鳥龍的體型非常小,擁有發展良好的翼,並有羽莖瘤(飛羽的附著點)的證據,它們即有可能有飛行能力。
小盜龍亞科是馳龍科中第二原始的演化支。小盜龍亞科包含許多最小的馳龍科恐龍,並顯示出樹棲生活的證據。所有馳龍科的皮膚痕跡證據都來自於小盜龍亞科,所有小盜龍亞科都覆蓋者羽毛並擁有發展良好的翼。如同半鳥亞科,有些物種可能擁有主動飛行能力。
伶盜龍亞科傳統上包含伶盜龍、恐爪龍、以及蜥鳥盜龍,而新發現的白魔龍可能屬於伶盜龍亞科,而蜥鳥盜龍的分類位置仍未確定。
馳龍亞科通常包含體型中到大的物種,並擁有箱形的頭顱骨;其他的亞科通常擁有狹窄的口鼻部。
以下馳龍科各屬的分類表是根據賽里諾(2005年)、森特(2004年)、馬克維奇等人(2005)、諾瑞爾等人(2006年)、以及特納(2007年)等人的研究而來;但不包含發現於英格蘭的偵察龍。
 大黑天神龍,一種原始馳龍科
大黑天神龍,一種原始馳龍科馳龍科 Dromaeosauridae
野蠻盜龍 Atrociraptor
似馳龍 Dromaeosauroides
欒川盜龍 Luanchuanraptor
大黑天神龍 Mahakala
火盜龍 Pyroraptor
烏奎洛龍 Unquillosaurus
 猶他盜龍,一種大型的馳龍亞科
猶他盜龍,一種大型的馳龍亞科瓦爾盜龍 Variraptor
馳龍亞科 Dromaeosaurinae
阿基里斯龍 Achillobator
惡靈龍 Adasaurus
馳龍 Dromaeosaurus
猶他盜龍 Utahraptor
 斑比盜龍,屬於小盜龍亞科
斑比盜龍,屬於小盜龍亞科小盜龍亞科 Microraptorinae
斑比盜龍 Bambiraptor
羽龍 Cryptovolans
纖細盜龍 Graciliraptor
西爪龍 Hesperonychus
小盜龍 Microraptor
中國鳥龍 Sinornithosaurus
 半鳥,屬於半鳥亞科
半鳥,屬於半鳥亞科半鳥亞科 Unenlagiinae
南方盜龍 Austroraptor
鷲龍 Buitreraptor
內烏肯盜龍 Neuquenraptor
脅空鳥龍 Rahonavis
Shanag
半鳥 Unenlagia
 伶盜龍,屬於伶盜龍亞科
伶盜龍,屬於伶盜龍亞科伶盜龍亞科 Velociraptorinae
恐爪龍 Deinonychus
偵察龍 Nuthetes
蜥鳥盜龍 Saurornitholestes
白魔龍 Tsaagan
伶盜龍 Velociraptor
系統發生學
在1998年,保羅·塞里諾(Paul Sereno)將馳龍科定義為一個演化支:包含馳龍,但不包含傷齒龍、似鳥龍、或家麻雀在內的最大天然族群演化支。馳龍科的不同亞科都曾被定義為演化支過,都常是以馳龍或其他次演化支作為定義基準點;舉例而言,馬克維奇將半鳥亞科定義為:在馳龍科中,所有親援關係接近半鳥,而離伶盜龍較遠的所有物種。小盜龍類(Microraptoria)是唯一不是亞科的次演化支,森特與其同事在建立時,沒有加上亞科的字尾-inae,以避免被誤認,並獨立於馳龍科之外。但塞里諾則提出不同的定義,並將小盜龍類改名為小盜龍亞科(Microraptorinae),歸類於馳龍科之內。但是,這個用法僅出現在他的TaxonSearch database網站,且未在正式文獻中出現過。
古生物學
 馳龍的腳部與第二趾爪
馳龍的腳部與第二趾爪掠食行為
關於馳龍科第二腳趾的鐮刀狀趾爪的功能,目前存在者不同的意見。在1969年,當約翰·奧斯特倫姆(John Ostrom)敘述恐爪龍的鐮刀狀趾爪時,他將這些趾爪解釋為類似刀片的砍傷用武器,相當類似劍齒虎的犬齒,恐爪龍使用強壯的趾爪來挖開獵物的身體。這個解釋被套用到所有馳龍科恐龍身上。然而,Manning等人認為這些趾爪並非當做鉤爪使用,並將它們重建為長橢圓形橫斷面,外圍包覆者角質層,而非原先認為的倒淚滴形橫斷面。在Manning的解釋中,第二趾爪可能在獵食大型獵食時,用來協助攀爬用,同時也是種刺傷武器。
約翰·奧斯特倫姆曾參考駝鳥與食火雞,發現這些鳥類會用較大型的第二趾爪攻擊獵物,進而產生恐爪龍會利用第二趾爪攻擊獵物的理論。食火雞的第二趾爪可長達12.5公分。奧斯特倫姆並引用一個1958年的鳥類論文,認為後肢與第二趾爪,可用來將獵物的身體撕開、將內臟挖出。在近年,Chhristopher P. Kofron研究241件食火雞的攻擊紀錄,發現總計有1個人類、2隻狗的被攻擊死亡案例,但沒有證據顯示食火雞會用第二趾爪將獵物的內臟挖出。相反地,食火雞將趾爪作為防衛武器,攻擊對它們產生威脅的動物、或威嚇對方。叫鶴也具有較大的第二趾爪,卻是用在撕裂小型獵物,以適合吞咽。
群體獵食
曾經在腱龍的化石附近發現一個小群體的恐爪龍化石;腱龍是種大型草食性鳥腳下目恐龍。這發現被解釋成馳龍科恐龍以有秩序群體獵食的證據,類似某些現代哺乳類。然而,並非所有古生物學家認為這證據具決定性,而後來的研究顯示恐爪龍較可能以無秩序的群體攻擊獵物。現代鳥類與鱷魚(馳龍科的近親們)顯示出很少協調的獵食行為;相反地,它們通常是獨自獵食,或是被屍體所吸引,同一物種之內可能存在者這兩種不同的方式。以印尼的科摩多龍來說,它們在群體進食的時候,最大型的個體會先進食,並攻擊企圖先進食的較小型個體;如果較小的個體死亡,將會遭到同類相食。當以這個現象來觀察原本被認為是群體獵食的馳龍科恐龍時,則呈現出同時符合科摩多龍與鱷魚的進食模式。這些地點所發現的恐爪龍骨骸都是來自於未成年體,而遺失的部份可能被其他恐爪龍所吞食,這不符合原先所認為的群體獵食行為。
在2007年,科學家們在中國山東省發現了第一個大量的馳龍科足跡化石。這個足跡化石可能是由體型接近阿基里斯龍的大型馳龍科恐龍所留下,而且是由六個相近大小的個體,沿者海岸共同前進。這些個體相距約一公尺,朝同一方向緩慢前進。研究人員提出,這個足跡化石可證明馳龍科的某些物種過者群體生活。雖然該足跡化石並不能明確指出馳龍科的獵食行為,但無法排除它們以群體獵食的可能性。此外,該發現還支持了馳龍科在行走時,第二腳趾後縮、離地的假設。
 顧氏小盜龍的羽毛痕跡。
顧氏小盜龍的羽毛痕跡。羽毛
有許多的證據顯示,馳龍科的身體覆蓋者羽毛。有些馳龍科化石的手部、手臂、尾巴,保存者長的正羽,而身體則保存者絨羽。另外有些化石,雖然沒有保存羽毛,但前臂骨頭上則有突起物,可能在生前是羽毛的附著處。整體而言,馳龍科的羽毛型態非常類似始祖鳥。
第一種被發現有明顯羽毛證據的馳龍科恐龍是中國鳥龍,由徐星等人於1999年在中國發現。許多其他的馳龍科化石曾發現羽毛覆蓋者身體,有些擁有完全發展的有羽毛翅膀。有些物種甚至顯示出後肢為第二對翼的證據,包括小盜龍與羽龍。因為羽毛痕跡只會在細緻的沉積層中保存下來,有些發現於粗糙岩層中的化石保有了羽莖瘤,羽莖瘤是鳥類的飛羽附著點。脅空鳥龍與伶盜龍的化石都發現了羽莖瘤,顯示它們的前肢具有羽毛。因此,大型、居住於地面的馳龍科恐龍極可能擁有羽毛,如同大部分的現代無法飛行鳥類仍保有它們的飛羽。雖然有些科學家主張,大型馳龍科失去了部份或全部的羽毛,但伶盜龍的羽莖瘤被視為是所有的馳龍科都具有羽毛的證據。
飛行與滑翔能力
目前只少有兩種馳龍科恐龍被認為具有飛行或滑翔能力,脅空鳥龍、顧氏小盜龍。脅空鳥龍最初被歸類於鳥類,後來的研究將其歸類於馳龍科。脅空鳥龍的長前肢被發現有羽莖瘤,羽莖瘤是飛羽的附著點,因此脅空鳥龍被認為可以動力飛行。從結構來看,脅空鳥龍的前肢比始祖鳥更有力,並附有強壯的韌帶,可以作出拍動的動作,展開飛行。路易斯·齊亞比(Luis Chiappe)提出脅空鳥龍具有這些適應特徵,可以飛行,但飛行能力較現代鳥類差。
顧氏小盜龍的前肢與後肢相當發達,科學家認為它們可以采四翼方式滑翔。在2005年,薩克·查特吉(Sankar Chatterjee)提出顧氏小盜龍的四肢類似雙翼飛機,但是采前後排列方式。查特吉認為它們在樹枝向下俯衝,以U字形的曲線滑翔,然後降落在地表或是其他的樹上;尾巴與後肢可用來控制方向與速度。此外,查特吉認為顧氏小盜龍有初步的飛行能力。
羽龍也被認為可能具有飛行或滑翔能力,但有科學家提出羽龍可能是小盜龍的異名。
大眾文化
1993年的電影《侏羅紀公園》(Jurassic Park)中所出現的伶盜龍,體型比該屬所發現最大的成員還大。羅伯特·巴克(Robert Bakker)說史蒂芬·史匹柏對於伶盜龍的體型改到失望,於是決定增加它們的體型;而猶他盜龍的體型較接近於電影中伶盜龍的體型。葛瑞格利·保羅(Gregory S. Paul)在他的書籍《Predatory Dinosaurs of the World》中,將恐爪龍列為伶盜龍的一種,並將該種命名為平衡伶盜龍(Velociraptor antirrhopus),這個分類法後來遭到嚴重地否決。麥克·克萊頓(Michael Crichton)將這兩個屬都寫入他的《侏羅紀公園》系列小說當中。在電影《侏羅紀公園》中出現的馳龍科恐龍,在當時是正確的描述,但現在看來則有許多方面是錯誤的,包含缺乏羽毛,但在《侏羅紀公園 III》(Jurassic Park III)中則將這個錯誤糾正回來。
外部連結
[1] 化石網 http://bbs.uua.cn/viewthread.php?tid=66670
