
形態構造
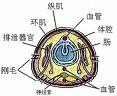
分節性身體由若干相似而沿其前後軸重複排列的體節或環節構成。相鄰體節間外部有環形溝或體環、內部以隔膜分界。蛭綱體節數恆定均為34節,其他環節動物 5~1000體節不等。身體分為頭部、軀幹部和肛部:頭部位於身體前端,多由口前葉和圍口節組成;軀幹部位於頭部和肛部之間。肛部具肛門,位於體之後端由 1節或若干節組成。環節動物的分節性既表現在體表,也表現在體內的重要器官(排泄器官、血管、生殖腺、神經節等)上。疣足和剛毛 除大部分蛭類外,多具上皮剛毛囊細胞產生的幾丁質剛毛。寡毛類剛毛較少,成對或成環直接著生於體壁;多毛類剛毛較多,且成束,多位於疣足葉上。疣足為軀幹部體節側生的葉片狀肉質突起,形態多樣,典型的疣足為雙葉型。自由運動的多毛類多具發達的疣足和剛毛,以利運動、游泳或錨定、攝食、感覺和呼吸。
體壁和消化管 體壁(外管)由角質層、單層柱狀上皮,不發達的基膜、外環肌層、內縱肌層和壁體腔膜組成完整或不完整的皮膚肉囊;消化管由髒體腔膜、縱肌層、環肌層和腸上皮組成。壁體腔膜和髒體腔膜間所包圍的腔隙即真體腔,經體腔管開口於體外。
體腔分室 環節動物發生時具若干分節排列的中胚層帶,由此發展為真體腔。相鄰的體腔由隔膜隔開,並經隔膜上的孔相通。有的種隔膜次生地部分或全部消失。體腔分室對環節動物的適應有重要作用。
分室的液壓骨骼 液壓骨骼的基本結構是個兩端封閉的筒,筒壁具環肌、縱肌層,筒內充滿液體。當縱肌收縮時,體腔液產生側向壓力,蟲體遂變寬縮短,疣足和剛毛錨在穴壁上;當環肌收縮時,體腔液產生從前到後的張力,蟲體變長且向前運動。由於環節動物體腔分室,液壓作用於局部體節,體形長短變化可限於一定體節,這就大大提高了液壓骨骼的效率。因此,環節動物爬行、游泳、穴居的能力甚強。
與其他低等蠕蟲(扁蟲、紐蟲、線蟲)相比,環節動物的器官系統更集中。體壁中胚層和真體腔的分室使環節動物強化了液壓骨骼,提高了環節動物的運動攝食力;髒壁中胚層分化為腸壁肌(加強了消化道的機械消化力和腸道的蠕動)以及腸血管叢(提高了對營養物質的吸收);閉管式循環系統以及具收縮力的背血管和心臟的分化,能使血液沿血管快速定向輸送營養物質、氣體和含氮廢物;原腎為兩端開口的後腎所替代加強了排泄功能;神經系統由腦(咽上神經節)、圍咽神經環、咽下神經節和各體節的神經節組成,鏈式神經系統的出現,使中樞神經的功能更加集中。
生殖和發生
寡毛綱、蛭綱雌雄同體,生殖腺分布於固定體節中,有與生殖關聯的複雜的附屬器官,性成熟後具特殊的腺狀體節(環帶),雄性先熟,異體交配受精,具卵繭,且直接發育;多數多毛綱動物常無固定的生殖腺,無環帶亦無交配現象、生殖產物直接排放在水中,受精卵經螺鏇卵裂發育成倒梨形的擔輪幼蟲。擔輪幼蟲多經過一段浮游期,逐漸變態沉落海底(底棲多毛類)。系統發生
環節動物一方面表現了原始的呼吸(多靠濕潤的體表或疣足葉、簡單的鰓)和運動機制,另一方面蠕蟲狀的外形雖與扁形動物和紐形動物相似,但其消化、循環和排泄系統趨於完善複雜、神經系統更集中因而又是比上述動物更進化的類群。通常認為,環節動物與軟體動物有共同祖先(擔輪祖體),分別向運動和不運動方向演化;此外,分節作為節肢動物的分部(頭、胸、腹部)的基礎,疣足作為節肢動物的附肢雛型,以及中樞神經系統上的相似性,又說明環節-節肢動物之間的相互關係。分類
通常分以下3綱:多毛綱 為環節動物門中最大的一綱。具發達的頭部和觸角、觸手、觸鬚、眼等感官;具疣足和成束分布的剛毛;具發達的體腔。多雌雄異體,發育多經擔輪幼蟲期。一般體長不超過10~50毫米。主要海生,大多底棲或浮游自由生活、極少數共棲或寄生。已知近萬種。
寡毛綱 頭部不發達,無疣足,剛毛直接生於體壁。具發達的體腔。成熟個體具環帶。雌雄同體,直接發育。多為陸生,亦具淡水生者。已知3100種,其中約200種為海生寡毛類,主要分布於潮間帶(特別是河口泥沙灘),體長多在20毫米以下,是小型底棲動物的主要組成成分。
蛭綱 體前、後兩端或僅後端具吸盤。體節數恆定為34節,且具次生環輪。無疣足,剛毛無或退化。體腔很不發達,為肌肉和結締組織所充填。具環帶。雌雄同體。體長3~250毫米。主要生活於淡水,陸生和海生者較少。已知約500種,其中75%為吸血或吸體液的外寄生者,寄主主要是脊椎動物,亦偶見於甲殼動物。
除上述3綱分類外,還有人將環節動物分為4綱〔多毛綱、寡毛綱、蛭蚓綱 (Branchiodellida)、蛭綱〕或5綱〔多毛綱、原環蟲綱(Archiannelida)、寡毛綱、蛭蚓綱、蛭綱;或多毛綱、原環蟲綱、吸口蟲綱、寡毛綱、蛭綱〕。有的學者根據環帶的有無分成環帶亞門 (Cli-tellata)和無環帶亞門 (Aclitellata)。80年代以來又有多毛綱、體蟲綱 (Aeolosomata)、環帶綱的分類方案。
多毛綱結構較其他各綱簡單、分化較少、生殖腺由體腔上皮形成,發育經擔輪幼蟲期等,通常被認為是較原始的類群。寡毛綱是由多毛綱祖先衍生而來並適於鑽土壤生活,而蛭綱則由寡毛綱進一步向寄生特化。原屬寡毛類的水生體蟲在許多方面(口前葉具纖毛的運動器官、腹神經末脫離上皮、雌雄同體、腎管兼做雄生殖管等)與沙間的多毛類相似,蛭綱中原始的棘蛭目體腔發達,前端具剛毛、具血管,與寡毛綱相近;寄生於淡水螯蝦,具15體節亦具隔膜的寡毛綱蛭蚓類,則因具吸盤、無剛毛等又與蛭綱關係密切。至於原環蟲類,許多學者認為是退性進化的多毛類,故多分散於多毛綱的各目和各科中。而吸口蟲是共棲或寄生於海百合或其他棘皮動物的體表上,因生活習性和體內外分節的特殊變化,屬於或從多毛綱中獨立出來。
意義
環節動物可提高土壤肥力,有利於改良土壤。可促進固體廢物還原;可供做餌料,增加動物蛋白質,可作為環境指示種;可用於醫療和入藥;另外,有的是有害的海洋污著生物。參考書目
吳寶鈴:環節動物多毛綱,《中國經濟動物志》,科學出版社,北京,1963。楊德漸、孫瑞平:《中國海習見多毛環節動物》,農業出版社,北京,1988。
配圖
 環節動物門
環節動物門