
物種起源
 腔腸動物的模式圖
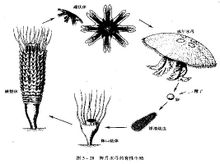
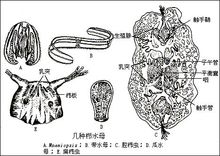
腔腸動物的模式圖腸動物是有了明確組織的真後生動物,因它們身體中央具有營消化、循環等功能的腔而得名。腔腸動物化石在前寒武紀即已出現。原水母綱以布魯克斯水母(圖1 幾種水母化石
)為代表,其體形變化大,一般呈星橢球狀,最早出現於寒武紀晚期至奧陶紀,在約1.5億年後的石炭紀又出現;分布於北美、歐洲和北非,在中國也有分布。側水母綱以狄更遜水母(圖1 幾種水母化石)為代表,其身體呈橢圓形鍾狀,兩側對稱,化石稀少,僅見於澳大利亞的下寒武統。因其具兩側對稱的特徵,被認為可能是蠕形動物或原始節肢動物。缽水母綱又稱真水母綱,以游水母(圖1幾種水母化石)為代表,傘狀,化石最早發現於美國科羅拉多大峽谷的前寒武系砂岩,此外,如蘇聯、瑞典、捷克斯洛伐克的中寒武統,加拿大和美國的上寒武統,德國的二疊系和侏羅系中也曾發現。
外形特徵
 腔腸動物的模式圖
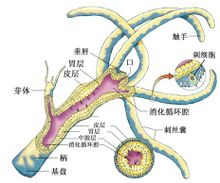
腔腸動物的模式圖刺胞動物體呈輻射或兩輻射對稱,僅具二胚層,是最原始的後生動
物。體壁由外胚層、內胚層和中膠層組成。內胚層圍成身體的整個內腔稱消化循環腔,腔腸一端為口,他端閉塞,無肛門。體壁中有刺細胞。腔腸動物的骨骼主要為外骨骼,具有支持和保護功能。多由幾丁質、角質和石灰質構成。在很多珊瑚蟲中具有骨針或骨軸,它們存在於中膠層,或突出於體表面。
生活習性
取食與消化
 腔腸動物門圖集
腔腸動物門圖集刺胞動物都是肉食性的,以浮游生物,小的甲殼類、多毛類甚至小的
魚類為食。由於食物的機械刺激和化學刺激,引起水螅類動物伸長觸手,並放出刺絲囊以纏繞、麻痹、毒殺捕獲物,再將食物送入口中。口區腺細胞分泌的粘液有利於食物的吞咽,食物進入胃腔後,胃層的腺細胞開始分泌蛋白酶,分解、消化食物使之形成許多多肽,同時在胃腔中由於營養肌肉細胞的鞭毛運動,食物得以混合與推動。經這種細胞外消化之後,開始細胞內的消化過程,營養肌肉細胞的偽足吞噬食物顆粒,在細胞內形成大量的食物泡,經過酸性及鹼性的化學過程之後,營養物質由細胞的擴散作用輸送到全身。缽水母類及珊瑚類胃腔結構比較複雜。缽水母的胃腔中有各種輻管及環管,胃囊中有內胚層起源的胃絲;珊瑚類的胃腔被許多隔膜分隔成許多小室,隔膜上有隔膜絲。胃絲及隔膜絲中含有大量的刺細胞及腺細胞,它們是將食物吞入胃腔之後才殺死及消化。消化後的營養物通過各種管道輸送到全身,未消化的食物殘渣仍由口排出。糖元及脂肪是腔腸動物的主要貯存物。刺胞動物中許多種類,特別是海洋中的造礁珊瑚類,體內均有共生的藻類,如動物黃藻、腰鞭毛藻之類。藻類能進行光合作用,產生甘油、脂肪、糖及脯氨酸等,並提供給刺胞動物作為其補充營養。這將在珊瑚綱中做進一步的敘述。
呼吸與排泄
 腔腸動物
腔腸動物刺胞動物沒有專門的呼吸及排泄器官,由於身體是由兩層細
胞圍繞胃循環腔所組成,並通過口使胃腔與外界相通,實際上體壁的兩層細胞均與外界環境接觸,所以呼吸與排泄作用可以由體壁細胞直接獨立進行。出、入口及胃腔的水流可以攜帶入新鮮的氧氣,並帶走代謝產物。刺胞動物的主要含氮廢物是氨。
生長繁殖
 腔腸動物的模式圖
腔腸動物的模式圖無性生殖及有性生殖在腔腸動物中都是很普遍的。無性生殖的主
要形式是出芽生殖,特別是在水螅型更為常見。例如水螅出芽時從身體的靠基部通過體壁及胃腔的向外突出,再長出觸手與口即形成芽體,以後芽體與母體分離即形成新的個體。但在藪枝螅(Obelia)所形成的芽體與母體不分離則形成了群體。其次,也可以通過分裂方式進行無性生殖,主要發生在水螅型,例如海葵可以縱分裂,缽水母的幼體以橫分裂進行無性生殖。此外水螅型一般具有很強的再生能力,例如,將水螅切成數段,條件合宜時每段都可以再生成一個新的個體,再生時口與反口端的極性不變,但口端的再生速度比反口端迅速。所以水螅型的再生現象也被認為是無性生殖的一種方式。有些生殖出現在多數水螅型及所有的水母型,除少數種類例如水螅為雌雄同體之外,絕大多數種類為雌雄異體(或異群體)。生殖細胞來源於間細胞,然後遷移到固定的位置上形成生殖腺。水螅蟲綱的動物生殖腺來源於表皮層,例如水螅,水螅水母的生殖腺位於放射管下或垂唇周圍,但都是來源於表皮層細胞。缽水母綱的生殖細胞起源於胃層位於胃囊低部。珊瑚綱的生殖細胞是由胃腔中的隔膜上發生,也是內胚層來源。腔腸動物只有生殖腺,沒有出現生殖導管及
生殖附屬腺,生殖細胞成熟之後由口排出,或由體壁破裂而釋放。受精作用依不同的種而異,或在體外海水中進行,或在垂管的表面,或在胃腔內生殖腺的部位進行。卵裂是完全的,形成中空的囊胚,經移入法或內陷法形成原腸胚,結果形成兩層細胞,即兩個胚層,內部成團的細胞為內胚層,將來形成成體的胃層,外表的一層為外胚層,將來形成成體的表皮層。實心的原腸胚迅速延長,體表出現纖毛,形成了自由游泳的浮浪幼蟲(planula)。浮浪幼蟲早期沒有口及胃腔,游泳一段時間之後,固著在水草、岩石或其他物體上發育成水螅型體,或再經過出芽生殖形成群體。淡水生活的水螅沒有幼蟲期,其受精卵直接發育。
概述
腔腸動物門(Coelenterata)又稱刺胞動物門。除極少數種類為淡水生活外,絕大多數種均為海洋生活,大多數在淺海,有些在深海,現存種類大約有11000種。
腔腸動物在動物界中的地位
腔腸動物是後生動物的開始, 所有其他後生動物都是經過這一階段發展起來的。
主要的特徵
 腔腸動物門
腔腸動物門(1)輻射對稱
輻射對稱即通過身體的中央軸有許多切面可以把身體分成相等的兩部分。
是一種原始的對稱方式,適應固著和漂浮生活,定向運動能力差。
一些種類(例如珊瑚綱)已經發展出兩輻射對稱。
兩種基本的結構類型,即水螅型(polyp)和水母型(medusa)。
(2)二胚層,原始消化腔
腔腸動物是真正的二胚層動物(內、外胚層) 。
在兩個胚層之間有中膠層。
體內的腔,即胚胎髮育中的原腸腔, 兼有消化和循環的作用,又稱消化循環腔。
有口無肛門。口為原口,有攝食和排遺的功能。
兼有細胞內消化和細胞外消化。
(3)有組織的分化
上皮組織占優勢,由它形成體內外表面,並分化為各種細胞,包括:上皮肌肉細胞(簡稱皮肌細胞)、腺細胞、間細胞、感覺細胞、刺細胞等。
(4)刺細胞
刺細胞是腔腸動物所特有的一種攻擊和防衛性細胞。
分布在外胚層細胞中,以口區、觸手上最多,但在缽水母綱及珊瑚綱的內胚層上也有大量分布,以幫助捕食。
每一個刺細胞有一囊狀的刺絲囊,囊中有毒液及盤繞的刺絲,刺細胞外側常有一刺針,能接受刺激,受刺激時刺絲連同毒液能立即射出,使對手麻醉或死亡。
(5)神經網
動物界裡最簡單最原始的神經系統。
沒有神經中樞,神經衝動的傳導一般是不定向的,被稱為擴散神經系統。
神經衝動的傳導速度比較慢。
(6)世代交替
世代交替指的是在動物的生活史中,無性世代與有性世代有規律地交替出現的現象。
水螅綱和缽水母綱的大部分種類存在世代交替現象。營固著生活的水螅體為無性世代;營自由生活的水母體為有性世代。水螅體以無性生殖(出芽或橫裂)產生水母型個體,水母體以有性生殖的方式產生水螅型個體。兩種世代有規律的相互交替。
珊瑚綱的全部種類以及水螅綱、缽水母綱的少數種類一生只有水螅型或者水母型,但是也具有有性生殖和無性生殖,只是二者並不交替進行。
(7)多態現象
水螅綱中有許多營群體生活的種類含有營養體與生殖體兩種形態與機能完全不同的個體,這種現象稱為二態現象。
群體中如果包括兩種以上不同形態與機能的個體,則稱為多態現象。
其生物學意義是群體中個體之間的勞動分工,通過群體中個體的形態分化來擔任不同的生理機能,使得群體成為一個有機的整體。
腔腸動物的主要分綱
 腔腸動物門
腔腸動物門水螅綱(hydrozoa)
缽水母綱(Scyphozoa)
珊瑚綱(Anthozoa)
水螅綱(hydrozoa)
本綱動物絕大多數海產,少數生活在淡水中。腔腸動物的淡水種類均屬於本綱。單體或群體生活。大部分種類生活史中有水螅型和水母型,或同時存在於群體中形成二態或多態,或交替出現形成世代交替;少數種類只存在水螅型或水母型。
常見種類有水螅(Hydra)、筒螅(Tubularia)、藪枝蟲(Obelia)、桃花水母(Craspedacusta)、鉤手水母(Gonionemus)、僧帽水母(Physalia)等。
缽水母綱(Scyphozoa)
本綱動物全部海產。生活史主要階段是單體水母,水母型構造比水螅水母複雜,水螅型不發達或完全消失,且常常以幼蟲的形式出現。
代表種類有各種大型水母,如:海月水母(Aurelia)、海蜇(Rhopilema)。
珊瑚綱(Anthozoa)
珊瑚綱是腔腸動物門最大的一個綱, 全部海產。
全部是水螅型的單體或群體動物,生活史中沒有水母型世代。珊瑚綱的水螅型結構較水螅綱複雜, 身體為兩輻射對稱。
常見種類如紅珊瑚(Corallium rubrum )、細指海葵(Metridium )、海仙人掌(Cavernularia )。
腔腸動物的系統發生
腔腸動物是真正多細胞動物的開始。
從其個體發育看,一般海產的腔腸動物都經過浮浪幼蟲階段,由此可推測:最原始的腔腸動物是能自由游泳、具纖毛、形狀象浮浪幼蟲的動物,即梅契尼柯夫假想的群體鞭毛蟲,細胞移入後形成原始二胚層動物(原始的水母型),發展成腔腸動物。
在現存的腔腸動物中,水螅綱無疑是最低等的一類,因為其水螅型與水母型的構構都比較簡單,生殖腺來自外胚層。缽水母綱水螅型退化,水母型發達,結構較複雜。珊瑚綱無水母型,只有結構複雜的水螅型。後兩綱的生殖腺又都來自內胚層,因此認為,缽水母綱和珊瑚綱可能起源於水螅綱,沿著不同的途徑發展而來的。
腔腸動物與人類的關係
有些種類有食用價值,有些可治療疾病。
毒素可作為新的藥物開發研究。
可用於地質學和油氣勘探。
珊瑚礁是全球生物多樣性最為豐富的生境之一,為其他動物的生存提供了多種環境。
仿生學,例如模仿水母的感覺器—觸手囊的風暴預報儀器,能提前15小時作出準確預報。
是生命科學研究的實驗材料,用於探討發育和進化等問題。
觀賞價值。
有些種類刺細胞分泌的毒液對人的危害較大,可造成嚴重創傷。
有些缽水母對漁業生產有害。
