
概述
 動物多樣性
動物多樣性動物界是多細胞真核生命體(真核生物域Eukaryota)中的一大類群。動物身體的基本形態會在它們發育時變得固定,通常是早在其胚胎髮育時,但也有些會在其稍後的生命中有個變態的過程。大多數動物是能動的,它們能自發且獨立地移動。
動物界是生態系統裡面的一個組成部分,絕大多數動物是消費者,間接利用太陽光的能源來生長。它們依靠其他生命體(如植物)作為其食糧。但也有少部分動物屬於分解者,以已經死亡的生物體(有機質)作為食糧(例如蚯蚓)。
動物界有著各種行為,塑造了生物圈的形態。這些行為可以看作是動物對刺激的反應。行為學是研究動物行為的科學。比較有名的行為理論是康納德·洛倫茨提出的本能理論。
大多數已知出現在化石中的動物門多是在5億4千萬年前的寒武紀大爆發時的海洋物種。以目前遺傳學的研究結果來看,動物的祖先應是來源於多種原生生物的集合,然後發生細胞分化,而不是來自一個多核原核生物。
截止到2005年,人類已知世界上的120萬種動物,其中有超過90萬種是昆蟲、甲殼類動物和蜘蛛類動物。
分類
動物學根據自然界動物的形態、身體內部構造、胚胎髮育的特點、生理習性、生活的地理環境等特徵,將特徵相同或相似的動物歸為同一類, 有脊索動物和無脊索動物兩大類。根據不同的準則,或稱為分類系統,動物會被不同地劃分。
按形態劃分
 動物
動物如果按形態學分類的話,動物首先按照組成的細胞數,分為單細胞動物(Protozoa)和多細胞動物(即後生動物Metazoa)兩種。前者所屬的動物有爭議,例如眼蟲,會因為其體內的葉綠體被歸入為植物。而在演化的過程中,多細胞生物中的細胞會因細胞分化而發展到不同的方向,行使不同的功能。
多細胞動物再被分為側生動物(Parazoa)和真後生動物(Eumetazoa)。前者包括海綿動物,扁盤動物和中生動物。這三種動物和真後生動物缺乏聯繫,組織分化程度低。接下來,真後生動物按照其身體對稱方式被分為輻射對稱動物和兩側對稱動物。前者包括刺胞動物門和櫛水母動物門。
然後將兩側對稱的動物按其體腔的有無,有的話是真是假,分為三類,即無體腔動物(Acoelomata),假體腔動物(Pseudocoelomata)和真體腔動物(Eucoelomata)。但是紐形動物門介乎於假體腔動物和真體腔動物之間,分類位置有疑問。無體腔動物的代表是扁形動物。假體腔動物的體腔並不是由中胚層包繞的,是原腸未完全退化的產物,代表動物是線蟲動物和輪形動物。真體腔動物的體腔是有中胚層包裹的。
真體腔動物接著按原腸孔(Blastoporus)的發展分為原口動物(Protostomia),後口動物(Deuterostomia)[21]和過渡類型觸手動物(Tentaculata)。後口動物的代表是棘皮動物和非“無脊椎動物”的脊索動物。過渡類型包括帚蟲動物,腕足動物和苔蘚動物三種。其他的真體腔動物都是原口動物,包括節肢動物,緩步動物,有爪動物,軟體動物,星蟲動物,螠蟲動物門和環節動物等。
這種分類有很大問題,比如紐形動物的“無家可歸”,而扁形動物,線蟲動物是原口動物,卻因為體腔不是“真體腔”而沒有“資格”去被歸類。觸手動物有很多後口動物的特徵,比如輻射卵裂,體腔是由內胚層內陷形成的中胚層包裹的。但是來自分子生物學的證據卻表明它們是原口動物。
根據18S rRNA比對結果劃分
遺傳學分類和按形態分類的出入在於兩側對稱動物中。如果按18srRNA序列比對的結果進行分類的話,兩側對稱動物首先按原腸孔的發展去向分為原口動物和後口動物。在形態學分類中的過渡類型觸手動物則被全部歸到原口動物中。
原口動物接著會按照蛻皮假說被分為兩種:蛻皮動物和冠輪動物。蛻皮動物的特徵是,這些動物在一種名叫蛻皮激素(Ecdyson)的作用下,會退去身體表面的角質層外皮。節肢動物,線形動物,緩步動物和有爪動物都屬蛻皮動物。冠輪動物的特徵是發育經過擔輪幼蟲階段(但有些動物發育過程中並不經歷幼蟲階段,很好的例子是蚯蚓)或是有觸手冠。軟體動物門(Mollusca)、環節動物門(Annelida)、紐形動物門(Nemertea)、星蟲動物門(Sipunculida)、螠蟲動物門(Echiura)、須腕動物門(Pogonophora),苔蘚動物門(Bryozoa)、內肛動物門(Entoprocta)、腕足動物門(Brachiopoda)和帚蟲動物門(Phoronida)都屬於這一冠輪動物。
這種分類方法沒有在形態分類學中出現的矛盾問題。不過卻還有一些地方需要進一步闡明,例如有爪動物的分類位置(位於蛻皮動物和冠輪動物之間)有爭議。
結構
除了少部份(如海綿)的例外,動物都有一個分化出分別組織的身體。這些組織包含肌肉(能收縮並控制身體的移動)和神經組織(傳遞與接收訊號)。一般也會有個內部的消化腔,和表皮連有一或兩個開口。有這些組織的動物被稱之為真後生動物。
所有的動物都有真核細胞,且被包在由膠原蛋白和具彈性的糖蛋白所組成的獨特細胞外網路之中。這些網路或許會鈣化以形成甲殼、骨頭和針骨等結構。在發育時會形成一個較可變動的架構,好讓細胞能移動且被重新組織,好使得複雜的結構變得可能。相對地,其他如植物和真菌等多細胞生物有被細胞壁固定住位置的細胞,所以以漸進的生長方式來發育。另外,動物細胞特有的還有如下幾種細胞間的結合:緊密接合、間隙接合和橋粒。
起源
 單細胞動物
單細胞動物 浮游雙瓣殼節肢動物
浮游雙瓣殼節肢動物動物界的歷史,就是動物起源、分化和進化的漫長曆程。是一個從單細胞到多細胞,從無脊椎到有脊椎,從低等到高等,從簡單到複雜的過程。
最早的單細胞的原生動物進化為多細胞的無脊椎動物,逐漸出現了海綿動物門、腔腸動物門、扁形動物門、紐形動物門、線形動物門、環節動物門、軟體動物門、節肢動物門、棘皮動物門。
由沒有脊椎的棘皮動物往前進化出現了脊椎動物,最早的脊椎動物是圓口綱,圓口綱在進化的過程中出現了上下頜、從水生到陸生。兩棲動物是最早登上陸地的脊椎動物。雖然兩棲動物已經能夠登上陸地,但它們仍然沒有完全擺脫水域環境的束縛。從原始的兩棲動物繼續進化,出現了爬行類。爬行動物可以在陸地上產卵、孵化,完全脫離了對水的依賴性,成為真正的陸生動物。爬行類及其以前的動物都屬於變溫動物,它們的身體會變得冰冷僵硬,這個時候它們不得不停止活動進入休眠狀態。然後爬行類動物進化為鳥類,成為了恆溫動物,不必進入休眠狀態,最後進化成胎生動物哺乳類動物,而人是哺乳類動物中最高級的動物。
進化
現代生物學研究表明,脊椎動物是由低等的無脊椎動物進化而來的,大致的進化過程是由與無脊椎的棘皮動物親緣關係密切的原始脊索動物中的半索動物或隱索動物到頭索動物或全索動物,最後可能由頭索動物或與其相近的種類進化出脊椎動物。由於原始脊索動物缺少硬體結構,不宜保存化石,因此至今發現的原始脊索動物的化石很少。
| 古生帶動物演化(時間單位:萬年) | |||||
570 | 500 | 435 | 395 | 345 | 280 |
寒武紀 | 奧陶紀 | 志留紀 | 泥盆紀 | 石炭紀 | 二疊紀 |
| 三葉蟲增多 | 珊瑚蟲、腕足動物、鸚鵡螺、筆石隨處可見 | 有鶚魚出現,海螺已存在 | 魚類增多,第一批兩棲動物出現 | 第一批爬行動物及第一批有翅昆蟲出現,兩棲類增多。 | 昆蟲種類增多,爬行動物占據陸地。 |
登入
 登入動物
登入動物脊椎動物由水生環境擴張到陸地環境,演化出陸生動物,這無疑是生命進化史上又一重大事件。因此,更確切地說,如果沒有動、植物實現登入的偉大進程,我們這個星球的生命還將停留在4億年前的古老而低級的階段。化石資料表明,兩棲類是最先由水中登上陸地生活的脊椎動物類群。
在兩棲動物生活史中,一方面仍保留著水中生活的階段,另一方面經過變態獲得生活在陸地上新的適應能力。然而,與植物登入相比,脊椎動物實現登入的時間要晚得多。最早的兩棲類化石發現在英格蘭老紅砂岩的頂部,地質時代屬於泥盆紀最末期(法門期末),距今大約有3.6億年。這是一類原始的、具有尾和迷齒、頭骨構造堅硬的早期兩棲類,即迷齒亞綱。魚石螈是這個古老類型的典型代表。
繁殖
幾乎所有的動物都會進行某種類型的有性生殖。成熟的個體是雙倍體或多倍體的。它們有一些特化的生殖細胞,行減數分裂以產生較小可遊動的精子或較大不可動的卵子。精子和卵子會結合成為受精卵,且發育成新的個體。受精卵一開始會發育成一個小球,稱之為囊胚,在此進行重整和分化。在海綿里,囊胚幼體會游到一個新的位置上並發育成一個新的海綿。而在其他大多數的類群中,囊胚則會進行更為複雜的重整。囊胚一開始會內套以形成具有消化腔的原腸胚和兩個各別的胚層-外胚層和內胚層。在大多數的情況下,還會有箇中胚層在兩者之間。這些胚層接著分化成各式組織和器官。
許多動物也能夠行無性生殖。這可能發生在孤雌生殖(成熟卵沒有經過交配而產生),或一些經由斷裂生殖。
門類
原生動物門
 胚胎幹細胞
胚胎幹細胞原生動物是最原始和最低等的動物類群。它們多數是單細胞生物,極少數是由幾個或多個細胞構成群體。原生動物的細胞是真核細胞,具有細胞的結構特徵,如細胞膜、細胞質、細胞核及其他細胞器。原生動物的細胞又是一個完整的生命體,具有多細胞生物表現出的生命功能,如從環境種吸取營養、呼吸、排泄、生殖,能夠對外界的刺激產生反應等。這些功能是細胞或由細胞特化而成的細胞器來完成的。
原生動物約有50000種,其中約有20000種為化石種。原生動物分為4個綱:鞭毛蟲綱(Mastigophora)、肉足蟲綱(Sarcodina)、孢子蟲綱(Sporozoa)、纖毛蟲綱(Ciliata)。
多孔動物門(Porifera)
 無脊索動物
無脊索動物多孔動物又稱海綿動物(Spongia),是多細胞動物中最原始最低等的類群,一般營固著生活。無固定體形,只有細胞分化,沒有真正的組織,如白枝海綿(Leucosolenia)。海綿動物的體壁由內、外兩層細胞和其間的中膠層(mesoglea)組成,有許多入水孔與體內特有的水溝系統相通。體壁內層主要是具鞭毛的領細胞(choanocyte),由於鞭毛的擺動,使水循著入水孔進入體內,以獲取食物和由入水孔排出代謝廢物。整個動物體壁大都由骨針和海綿絲支持。水溝系是海綿動物所特有的,它在固著動物生活的取食、排泄等方面起著重要作用。多孔動物雖是多細胞動物中最原始的類群,但它們卻是動物系統進化的一個側支。
腔腸動物門(Coelenterata)
腔腸動物是真正的二胚層多細胞後生動物,如水螅(Hydra)。
1. 腔腸動物的主要特徵
第一,輻射對稱。
第二,有兩胚層。
 水螅
水螅第三,有組織分化。
2. 腔腸動物的生殖方式
包括無性生殖和有性生殖。無性生殖最普遍的是出芽生殖。有性生殖在其生殖期產生精巢和卵巢,雌雄同體。藪枝螅有明顯的世代交替,這是動物中少有的特徵。
3. 腔腸動物的分類
腔腸動物約9000種以上,分為三個綱:
水螅綱(Hydrozoa),單個或群體,生活史中多有水螅型和水母型,如水螅和藪枝螅(Obelia)。
缽水母綱(Scyphozoa),多為大型水母,如海月水母(Aurelia)、海蜇(Rhopilemaesculentum)。
珊瑚綱(Anthozoa),只有水螅型世代,大多數具有發達的石灰質骨骼,如海葵(Metridium),各種石珊瑚。
扁形動物門(Platyhelminthes)
⒈扁形動物的進化特徵
第一,兩側對稱。第二,三胚層出現。第三,器官系統分化。
⒉扁形動物分類
扁形動物約有7000種,分3個綱:渦蟲綱(Turbellaria)、吸蟲綱(Trematoda)和絛蟲綱(Cestoioda)。
線形動物門(Nemathelminthes)
線形動物是一大類群,在動物系統進化上,出現了一個進步性的特徵,即假體腔(primarycoelom),如人蛔蟲(Ascarislumbricoides)。人蛔蟲營腸內寄生,身體表面覆蓋著角質層,體內器官退化,生殖器官特別發達,雌雄異體。
環節動物門(Annelida)
 蚯蚓
蚯蚓環節動物包括各種蚯蚓、沙蠶、螞蟥等。環節動物在動物系統進化上具有重要意義。
⒈環節動物的進化特徵
第一,身體出現分節現象(metamerism),這是高等無脊椎動物的一個重要標誌。然而環節動物的分節,仍屬原始分節現象。
 蚯蚓
蚯蚓第二,環節動物出現了真體腔(coelom),真體腔是由中胚層分化出來的,由壁體腔膜和髒體腔膜圍繞而成,因而體壁和腸壁都有發達的肌肉。
第三,環節動物器官系統較完善。
⒉環節動物分類
多毛綱(Polychaeta),如沙蠶,海產,有發達的頭部和疣足,雌雄異體,發育經過擔輪幼蟲期。
寡毛綱(Oligochaeta),如環毛蚓,無疣足而有剛毛,有生殖帶,雌雄同體,發育經過擔輪幼蟲期。
蛭綱(Hirudinca),如蛭,亦叫螞蟥,無疣足無剛毛,體節數目固定,身體前後端有吸盤,營寄生或半寄生生活。
軟體動物門(Mollusca)
軟體動物的數量僅次於節肢動物,約有10萬種,為動物界中第二大類群,包括各種螺類、蚌類、烏賊、章魚等。
軟體動物雖然在外形上差別很大,但主要形態結構基本相同。內臟結構較環節動物複雜,但在進化特徵上沒有明顯的進步。從胚胎髮育角度分析,軟體動物也經過擔輪幼蟲期,說明軟體動物和環節動物兩者親緣關係極為密切。
 章魚
章魚 軟體動物經濟意義較大,絕大多數種類可供食用。有的軟體動物可以作藥用,如鮑魚的貝殼是中藥石決明,烏賊的內骨骼是海螵蛸。珍珠既可入藥,又是名貴裝飾品。有些軟體動物對人類有害,鑿船貝(teredo)嚴重危害海中木船和木質建築,蝸牛是田園、果樹和農作物害蟲,釘螺等是寄生蟲的中間寄主。
節肢動物門(Arthropoda)
節肢動物是動物界中數量最多的一類無脊椎動物,占動物界動物總數150萬種的84%,約126萬種,其中昆蟲又占節肢動物的94%,約118萬種,故以昆蟲的特徵為例介紹節肢動物的主要特徵。全身包被外骨骼身體異律分節,有靈活的附肢、以及發達的肌肉。具有混合體腔和開管式循環系統具備氣管等空氣呼吸器,能高效地進行呼吸完全適應於陸上生活。
⒈節肢動物(昆蟲)的特徵
 狼蛛
狼蛛第一,異律分節的高度發展。
第二,體被幾丁質蛋白質複合體的外骨骼。
第三,附肢分節並有關節。
第四,橫紋肌發達,能迅速收縮。
第五,開管式血液循環。
第六,體壁內陷形成氣管作為呼吸器官。
第七,排泄系統由腸壁向外突起而形成。
第八,神經系統和感覺器官發達。
第九,絕大多數屬雌雄異體,體內受精。
⒉節肢動物分類
節肢動物分為3個亞門7綱:
有鰓亞門(Branchiata),大部分水生,少部分陸生,用鰓呼吸。包括三葉蟲綱和甲殼綱。三葉蟲綱已滅絕。
有螯亞門(Chelicerata),大部分陸生,少數水生。包括肢口綱(Merostomata),海產。
有氣管亞門(Tracheata),大部分陸生,少數水生。主要包括珠形綱、多足綱和昆蟲綱。
棘皮動物門(Echinodermata)
棘皮動物門包括海星、海膽、海參、海百合等動物,這是一個相當特殊的類群,雖然數量不多,但在進化上有其獨特的意義。
⒈進化特徵
 海膽
海膽第一,首次出現由中胚層起源的真(內)骨。
第二,在胚胎髮育過程中,胚胎期的胚孔形成肛門,在胚孔的另一端形成口,故稱後口。
第三,真體腔發達,且一部分真體腔形成特殊的水管系統。
第四,成體是五輻射對稱,器官也是按該體制排列,但棘皮動物幼體是兩側對稱,也就是說棘皮動物的五輻射對稱是在進化過程中適應海棲生活形成的次生現象,故和腔腸動物的原始輻射對稱是完全不同的。
第五,多數為雌雄異體,少數為雌雄同體,有胚胎髮育過程中經過的幼蟲階段與半索動物的幼蟲相似,表明棘皮動物是向著脊索動物進化過程中的無脊索動物。
2.外部形態:
 海百合
海百合大多數為五輻射對稱,但是次生形成的,由兩側對稱的幼體發展而來。口面:體盤中央為口,各腕腹側中央有一條自口伸向腕端部的步帶溝。反口面:近體盤中央處有肛門。兩腕間為間步帶區,一間步帶區有一篩板。各腕的基部兩側各具一對生殖孔,腕頂端有紅色眼點。
3.分類
約有5300種,全部為海中底棲生物,分為海星綱(Asteroidea);海蛇尾綱(Ophiuroidea),腕細長者如陽遂足(Amphiura);海膽綱(Echinoidea),無腕,骨骼形成堅固的球形,如馬糞海膽(Hemicentrotus
pulcherrimus)等;海百合綱(Crinoidea),以柄固著營永久性或暫時固著生活,有5或5倍數的輻射羽狀腕,如小羽枝(Oligometrachinensis)。此外還有海參綱(Holothuroidea),沒有互相關聯的骨板,骨片小,分散在皮膚內,長筒形,口周圍管足變成觸手。
半索動物門(Hemichordata)
是動物界中數量較少的類群之一,代表動物柱頭蟲。它是動物系統上一個獨特的門類。
⒈進化特徵
第一,有鰓裂,鰓裂(gillslits)是從本門動物開始出現的較高等的進化特徵。
第二,背神經索中有狹窄的空隙出現,可能與高等脊索動物的中空背神經管同源。
第三,有口索(stomochord),口腔中前端背面伸至吻基部的一條短盲管。大量研究表明,口索和脊索既不同功也不同源,故此類動物為半索動物。
⒉分類地位
雖然柱頭蟲有許多脊索動物的特徵,但它仍是無脊索動物,且通過對生化成分測定,與棘皮動物親緣關係更近,故它是無脊索動物向脊索動物過渡的無脊索動物。
脊索動物門(Chordata)
脊索動物是動物界中最高等的一類動物,脊索動物的共同特徵有:
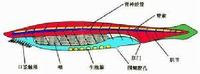 脊索動物
脊索動物第一,脊索(notochord)。脊索是動物身體背部起支持作用的一條棒狀支柱,位於消化道背面,神經管的腹面,有彈性而不分節。脊索內部含富有液泡的細胞,外面圍有厚的、由結締組織構成的脊索鞘。
第二,背神經管(dorsaltubularnervecord)。中空的背神經管位於脊索的背面,起源於外胚層。脊椎動物的神經管前部膨大形成腦,腦以後的神經管發育成脊髓。神經管內腔在成體腦中形成腦室,在脊髓中成為中管。
第三,鰓裂(gillslits)。鰓裂是咽部兩側一系列成對的裂縫,與外界相通,是水生脊索動物的呼吸器官。
第四,心臟在腹面,閉館式循環系統(尾索動物除外)。無脊椎動物若有心臟則在背面。
脊索動物現在約有50000種,分為尾索動物亞門、頭索動物亞門和脊椎動物亞門。尾索動物和頭索動物通稱原索動物(Protochordata)。
分類系統
 哺乳類
哺乳類分類系統是階元系統,通常包括7個主要級別:種、屬、科、目、綱、門、界。種(物種)是基本單元,近緣的種歸合為屬,近緣的屬歸合為科,科隸於目,目隸於綱,綱隸於門,門隸於界。隨著研究的進展,分類層次不斷增加,單元上下可以附加次生單元,如總綱(超綱)、亞綱、次綱、總目(超目)、亞目、次目、總科(超科)、亞科等等。
還可增設新的單元,如股、群、族、組等等,其中最常設的是族,介於亞科和屬之間。通常種下分類,動物只設亞種單元。
