敘述
特暴龍是最大型的暴龍科動物之一,但略小於暴龍。已知最大型的個體身長10到12米,頭部離地面約5米。目前還沒有完全成長個體的體重數值,但它們一般被認為略輕於暴龍。目前已知最大型的特暴龍顱骨長度高過1.3米,小於暴龍,但大於其他的暴龍科。如同暴龍,特暴龍的顱骨高大,前段狹窄。顱骨後段擴張幅度不大,意味者特暴龍的眼睛不是直接朝向前方,所以它們缺乏暴龍擁有的立體視覺。顱骨的大型洞孔可減輕重量。嘴部有60到64顆牙齒,略少於暴龍,但大於其他體型較小的暴龍科,例如蛇發女怪龍與分支龍。大部分的牙齒橫剖面為橢圓形,而前上頜骨的牙齒橫剖面為D形。暴龍科都具有這種異型齒特徵。上頜骨的牙齒最長,齒冠長達85公厘。特暴龍與分支龍的下頜外側各有一道棱脊,從隅骨延伸到齒骨後方,形成相扣的結構。其他暴龍科動物缺乏這道棱脊,因此下頜更為靈活。
暴龍科的身體外形差異不大。特暴龍的頸部為S狀彎曲,其餘的脊柱與長尾巴,與地面保持者水平的姿態。特暴龍有者暴龍科中最小的前肢/身體比例,有兩根迷你的手指。每個手掌有兩根手指,某些標本被發現有退化的第三指,沒有指爪,其他暴龍科近親也有發現這點特徵。特暴龍的第二指長度小於第一指的兩倍,而其他暴龍科的第二指長度約是第一指的兩倍。另外,與其他暴龍科相比,特暴龍的第三指比例較小。某些暴龍科的第三指長於第一指,例如:亞伯達龍、懼龍;而特暴龍的第三指短於第一指。
特暴龍的後肢長而粗厚,將身體支撐為二足的步態,上有三根腳趾。長而重的尾巴可以平衡頭部與胸部的重量,將重心保持在臀部。
分類系統
特暴龍屬於暴龍科的暴龍亞科。該亞科還包含較早期的懼龍、較晚期的暴龍,都發現於北美洲,可能還有蒙古的分支龍。暴龍亞科包含親緣關係較接近暴龍,而離艾伯塔龍較遠的物種;與艾伯塔龍亞科相比,暴龍亞科的體格較重型,頭顱骨的比例較大,以及較長的股骨。勇士特暴龍最初被視為暴龍的一個種,某些近年的分類也支持這個說法。其他的科學家則將它們列為獨立的屬,並為暴龍的姐妹分類單元。在2003年,一個親緣分支分類法研究提出分支龍是特暴龍的最近親,因為它們具有其他暴龍亞科沒有的頭部特徵。如果屬實,將排除特暴龍是暴龍的一個異名的可能性,並顯示暴龍亞科在北美洲與亞洲演化出個別的支系。分支龍的兩個標本具有幼年體的特徵,但牙齒數量較多,約76到78顆,而且口鼻部上面有獨特的低矮骨質瘤,因此並非特暴龍的幼年體。
發現與命名
雖然已經發現許多特暴龍標本,在1986年以前很少已確定的明確資料,但特暴龍被推論與其他暴龍科恐龍有許多共同特徵。這些相近的特徵讓某些科學家提出當時的北美洲與歐亞大陸之間有可能的連結,也許是陸橋。在1946年,一個蘇聯與蒙古挖掘團隊在蒙古南戈壁省的耐梅蓋特組發現一個大型頭顱骨與一些脊椎骨,屬於一個大型的獸腳類恐龍。在1955年,蘇聯古生物學家葉甫根尼·馬列夫(Evgeny Maleev)將這正模標本(PIN 551-1)建立為暴龍的一個種,勇士暴龍(Tyrannosaurus bataar)。種名баатар/baatar在蒙古語中意為“勇士”,但被誤拼為bataar。
同年,馬列夫將3個獸腳類的化石敘述並命名,三者都是頭顱骨,外加部分身體,全是同一個挖掘團隊在1948到49年間發現的。第一個(編號PIN 551-2)被建立為新屬,埃夫雷莫夫特暴龍(T. efremovi),ταρβο?/tarbos在希臘文文中意為“恐怖的”、“敬畏的”,而σαυρο?/saurus意為“蜥蜴”,種名是以蘇聯古生物學家兼科幻小說作者伊凡·埃夫雷莫夫(Ivan Yefremov)為名。另外兩個(編號PIN 553-1與PIN 552-2)被歸類於北美洲的蛇發女怪龍,G. lancinator與G. novojilovi。這三個標本都小於1946年發現的標本。
在1965年,蘇聯古生物學家阿納托利·康斯坦丁諾維奇·羅特傑斯特文斯基(Anatoly Konstantinovich Rozhdestvensky)認為馬列夫所發現的標本,其實是同種動物的不同生長階段,而且不同於北美洲的暴龍。他將四個標本與新發現的化石,統合為勇士特暴龍(T. bataar)。後來的研究人員都同意羅特傑斯特文斯基的分類,包含馬列夫在內,但少數科學家使用埃夫雷莫夫特暴龍,而非勇士特暴龍。在1992年,美國古生物學家肯尼思·卡彭特(Kenneth Carpenter)重新檢驗這些化石,他根據頭顱骨的相似處,提出這些化石大部分屬於暴龍,並將勇士特暴龍改回勇士暴龍;而蛇發女怪龍的G. novojilovi是個較小型的暴龍科動物,另外建立為馬列夫龍(Maleevosaurus novojilovi)。在1995年,喬治·奧利舍夫斯基(George Olshevsky)建立了勇士成吉思汗龍(Jenghizkhan bataar),取代勇士特暴龍,並以成吉思汗為名;他並提出埃夫雷莫夫特暴龍、馬列夫龍、勇士成吉思汗龍是三個獨立的屬,都生存於相同時期的耐梅蓋特組。在1999年,一個研究提出馬列夫龍是特暴龍的幼年個體。自從1999年以來,所有的研究認為這些化石皆為同種動物,勇士特暴龍或是勇士暴龍。
自從蘇聯與蒙古的挖掘團隊在40年代的挖掘過後,一個波蘭與蒙古的挖掘團隊再度回到戈壁沙漠挖掘,從1963年持續到1971年,發現了許多新的化石,並在耐梅蓋特組發現了特暴龍的新標本。在1993年到1998年,日本與蒙古的挖掘團隊,以及21世紀初,加拿大古生物學家菲力·柯爾(Phil Currie)的挖掘小組,也發現了許多特暴龍化石。目前已發現了至少30個標本,包含超過15個頭顱骨,與數個完整的顱後身體骨骼。
特暴龍的化石發現於中國與蒙古國的戈壁沙漠,兩國都禁止化石出口,但某些化石落入私人收藏家手中。
可能異名
在60年代中期,中國古生物學家在中國新疆吐魯番盆地的蘇巴什組(晚白堊紀)發現了一個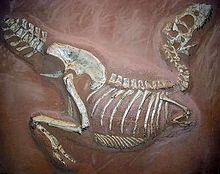
在1976年,Sergei Kurzanov命名了分支龍,是種發現於蒙古的暴龍科動物,年代稍早於特暴龍。數個研究指出分支龍是特暴龍的近親。Hurum與Sabath在2003年證實特暴龍是分支龍的近親,而非暴龍的一個種。這是根據暴龍科恐龍起源於亞洲,而且遷徙到北美洲的假設。其他的研究人員也支持這個意見。分支龍被認為是個成年個體,但它的長、低矮頭部卻是幼年暴龍科的特徵。基於此一特徵,柯爾推測分支龍是特暴龍的一個幼年體。但柯爾也認為,分支龍較多的牙齒數量,以及口鼻部上方的低矮瘤狀物,有其他可能的存在。
古生物學
如同其他大型暴龍科恐龍,特暴龍已發現了大量且保存良好的化石。事實上,耐梅蓋特組的化石大約有1/4的化石是特暴龍。雖然特暴龍不像北美洲的暴龍科一樣地經過徹底的研究,可些化石以足夠科學家們做出有限的生物學研究。頭部力學
在2003年,特暴龍的顱骨首次經過完整的研究。科學家們發現特暴龍與北美洲暴龍科之間有幾個顯著的差異。這些差異與顱骨在咬合時,力量的傳遞有關。當特暴龍的上頜咬住物體時,力量從上頜骨傳遞到上頜周遭的顱骨。而北美洲暴龍科咬合時,力量從上頜骨傳遞到口鼻部上方的固定鼻骨,然後再經由鼻骨與淚骨間的骨質連結,傳遞到淚骨。特暴龍的鼻骨與淚骨間缺乏骨質的連結。但上頜骨的後方有個大型突起,楔合入

另一個主要的差別則是堅固的下頜。許多獸腳類恐龍,包括北美洲暴龍科在內,下頜的齒骨與後面骨頭間有靈活的關節。特暴龍的隅骨側邊棱脊連線者齒骨後方的方形突,使它們的下頜無法靈活外內扳動。
有些科學家提出假設,認為特暴龍的堅硬頭部是種適應演化,用來獵殺耐梅蓋特組的大型蜥腳類恐龍,泰坦巨龍類,因為晚白堊紀的北美洲並沒有如此巨大的恐龍存在。這種頭部力學的差異也影響了暴龍科的種系發生學。同樣發現於蒙古的分支龍,也具有類似特暴龍的頭部力學特徵,這顯示暴龍並非特暴龍的最近親。特暴龍與暴龍的相似處可能導因於它們巨大的體型,是平行演化的結果。
腦部結構
一個在1948年發現的顱骨,一度被歸類於蛇發女怪龍的G. lancinator,因為具有顱腔,有助於科學家了解特暴龍的腦部結構。在1965年,馬列夫製作了一個石膏顱腔模型,並做了初步的腦部形狀檢查。在2005年,謝爾蓋·薩伐黎耶夫(Sergei V. Saveliev)製作了一個聚氨酯顱腔模型,並做了更詳細的腦部結構與功能研究。根據暴龍的腦部結構研究,它們兩者的腦部結構相似,只有某些腦神經根部的位置不一樣,包含三叉神經與副神經。暴龍科的腦部較類似鱷魚與其他爬蟲類,而較不類似鳥類。特暴龍的腦部體積為184立方厘米。腦部的大型嗅球、末端神經、嗅神經,顯示特暴龍具有靈敏的嗅覺,這點如同暴龍。犁鼻球大型,而且與嗅球分離,顯示可以感應費洛蒙的犁鼻器發展良好。這個特徵顯示特暴龍可能具有複雜的求偶行為。聽神經也很大,顯示它們的聽力很好,可能用在聲音的溝通與警告上。聽神經連線者發展良好的前庭系統,顯示它們的平衡感與協調性很好。相反地,與視力有關的腦部區域與神經較小。爬蟲類的中腦頂蓋連線控制眼球的視神經與動眼神經,是用來處理視力訊息,但特暴龍的中腦頂蓋非常小。暴龍的眼睛朝前,因此具有一定程度的立體視覺;但特暴龍的顱骨狹窄,眼睛朝向兩側,如同典型的暴龍科動物。這些特徵顯示特暴龍較依靠嗅覺與聽覺,而非視覺。
生長模式
特暴龍的大部分化石,是成年或亞成年個體,很少發現幼年個體的化石。在2006年發現的一個幼年個體身體骨骼,帶有完整的頭顱骨,頭部長度為29厘米,使得古生物學家可以研究特暴龍的生長模式。這個幼年個體化石,死亡時的年齡大約是2到3歲。與成年個體相比,這個幼年頭顱骨的結構虛弱,牙齒較細,顯示特暴龍的幼年個體、成年個體占據不同的生態位,以免競爭相同食物來源。病徵
在2001年,布魯斯·羅斯柴爾德(Bruce Rothschild)等人發表一份獸腳類恐龍的壓力性骨折與肌腱撕裂傷研究,並研究它們的行為模式。壓力性骨折較常導因於習慣性動作,較少來自於外力衝擊。他們研究18個特暴龍的腳掌骨骼,沒有發現壓力性骨折的跡象;而在10個被研究的手掌骨頭中,其中一個手部骨頭有發現壓力性骨折的跡象。腳部骨頭的壓力性骨折,可以歸咎於奔跑、長途遷徙。手部骨頭的壓力性骨折,極可能導因於捕抓獵物而受傷。這些壓力性骨折、肌腱撕裂傷跡象,顯示特暴龍常因為捕抓獵物而受傷,而非吞食屍體。古生態學
大部分的特暴龍化石出土於蒙古南部的耐梅蓋特組。這個地層組沒有經過放射性同位素計年,但根據該地化石記錄中的動物群,耐梅蓋特組的年代可能為晚白堊紀的馬斯特里赫特階。馬斯特里赫特階約為7,000萬年前。而發現鄯善龍的新疆蘇巴什組,地質年代也為馬斯特里赫特階。耐梅蓋特組保存了大型河道與土壤沉積物,顯示該地與更古老的巴魯恩戈約特組、德加多克塔組相比,曾經有過更潮濕的氣候。但硝石沉積物則指出該地曾有過周期性的旱季。沉積物分布在過去的大型河道與泛濫平原上。該地偶爾可發現軟體動物化石,以及其他多樣性的水生動物化石,例如魚類與烏龜。鱷類化石也被發現,例如數種沙漠鱷,沙漠鱷的牙齒適合壓碎貝殼。哺乳類的化石相當少見,鳥類化石的數量較多,包含反鳥亞綱的Gurilynia、黃昏鳥目的尤氏鳥(Judinornis)、以及雁形目的早期物種Teviornis。耐梅蓋特組所發現的恐龍,則包含:甲龍科的多智龍、厚頭龍下目的平頭龍與傾頭龍。該地層目前所發現最大型的掠食動物是特暴龍,它們極可能以大型鴨嘴龍類為食,例如櫛龍與巴思缽氏龍,或是蜥腳類恐龍,例如納摩蓋吐龍與後凹尾龍。成年特暴龍可能與其他小型獸腳類恐龍有少許競爭,例如傷齒龍科的無聊龍、鴕鳥龍、蜥鳥龍,以及偷蛋龍下目的單足龍、耐梅蓋特母龍(Nemegtomaia)、瑞欽龍,或者還有小掠龍,一種有時被認為是基底暴龍超科的恐龍。其他的獸腳類恐龍,包含:巨大的鐮刀龍、似鳥龍下目的似鵝龍、似雞龍、恐手龍,鐮刀龍可能是草食性動物,而上述似鳥龍類恐龍可能為雜食性動物,以小型動物為食,不會跟特暴龍競爭食物。如同其他大型暴龍科與現代的科莫多龍,特暴龍的幼年與近成年個體的身上發現過有齒痕,符合成年個體或其他較小型獸腳類恐龍的牙齒。
大眾文化
- 在澳洲維多利亞州的墨爾本博物館可看到一個特暴龍骨骸模型。雪梨麥覺理大學總圖書館入口處也有一個展示中的特暴龍標本。
- 特暴龍出現在BBC的2005年電視節目《恐龍凶面目》(The Truth About Killer Dinosaurs)第二集,以及《與恐龍共舞》(Walking with Dinosaurs)的特別節目《鐮刀龍探秘》(The Giant Claw)。
