
定義
經過兩性生殖細胞的結合而產生的受精卵,這種方式叫做有性生殖。有性生殖是通過生殖細胞結合的生殖方式。通常生物的生活周期中包括二倍體時期與單倍體時期的交替。二倍體細胞借減數分裂產生單倍體細胞(雌雄配子或卵和精子);單倍體細胞通過受精(核融合)形成新的二倍體細胞。這種有配子融合過程的有性生殖稱為融合生殖。某些生物的配子可不經融合而單獨發育為新個體,稱單性生殖。起源
 動物的有性生殖
動物的有性生殖雌雄兩個親體的性細胞(又稱配子)結合而發育成的一個新個體,此為兩性生殖,這在生物學上具有重要意義。新個體繼承了雙親的遺傳性狀,保持了種族的遺傳特性。大多數動物都進行有性生殖。絕大多數無脊椎動物為卵生,即雌性個體產生卵,卵與精子結合,形成幼體,幼體經生長發育成為成體。少數為卵胎生,即卵與精子結合後留在雌性體內發育,其所需營養由卵來供應,形成幼體後,產出體外,如田螺生下的為幼螺。也有的稱胎生,即雌體產生的卵受精後留在體內,並吸收母體的營養而發育成幼體,然後產出體外,如蚜蟲生下的小蚜蟲即屬此。在低等動物類群中,雌雄兩個親體所產生的性細胞,其大小、形態等基本相似,辨別不出雌雄,稱為同形配子;但多數動物的配子其形態相異,稱異形配子。如雄配子的精子多似蝌蚪狀,雌配子的卵則呈球形。有些無脊椎動物是雌雄同體(hermaphrodatism),即一個個體既可產生精子,又可產生卵。進行生殖時,也是精子與卵結合,不過有時是自己本身的精子和卵子結合,稱自體受精,或者自己本身的卵與其他個體的精子結合稱異體受精。如渦蟲、蚯蚓、蛭類等都是雌雄同體的動物。
主要方式
 有性生殖
有性生殖接合生殖細菌的接合生殖兩個菌體通過暫時形成的原生質橋單向的轉移遺傳信息。供體(雄體的部分染色體)可以轉移到受體(雌體)的細胞中並導致基因重組。這是最原始的接合生殖。
原生動物的接合生殖多見於纖毛蟲類,按接合的雙方,即接合子的形態又可分為兩類:
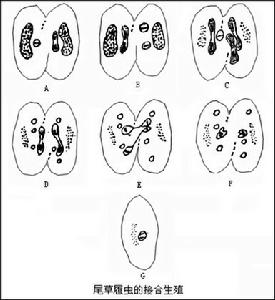
1、同配接合:接合子的形態相同。接合時雙方暫時融合,小核在減數分裂後進行交換,相互受精後分開,如尾草履蟲。
接合雙方緊靠在一起,口部融合,然後大核消失小核分裂二次,成4個,其中3個退化,一個再經一次分裂成為一個動核和一個靜核。此時接合個體互相交換動核,然後靜核與換來的動核融合。接著蟲體分開,每個個體的融合核分裂三次形成8個核,其中4個成大核,3個退化。此後大核不分裂,剩下的小核與蟲體同時分裂兩次而成為4個新的子體。
2、異配接合:見於緣毛目類纖毛蟲。在進行接合生殖前,蟲體先經一次不均等分裂,除小核外大核和蟲體都分成大小兩部分,成為大接合子和小接合子,前者固著,後者自由游泳。小接合子找到大接合子後即牢固附著在其上開始接合。在接合過程中,合子核只在大接合子中形成,小接合子為大接合子吸收。如鍾蟲。
配子生殖配子是由營養個體所產生的生殖細胞,需兩兩配合後才能繼續其生活史,如在一定時間內找不到適當的配子便死亡。按配子的大小,形狀和性表現可分為三種類型:
同配生殖配子的形態和機能完全相同,沒有性的區分。例如衣藻屬中的大多數種類。
異配生殖有兩種類型:①生理的異配生殖,參加結合的配子形態上並無區別,但交配型不同,在相同交配型的配子間不發生結合,只有不同交配型的配子才能結合,且具有種的特異性。如衣藻屬中的少數種類。這是異配生殖中最原始的類型。②形態的異配生殖,參加結合的配子形狀相同,但大小和性表現不同。大的不大活潑為雌配子,小的活潑,為雄配子,這說明已開始了性在形態上的分化。
配子生殖的進化趨勢是由同配到異配,最後發展為卵配生殖。在原生動物和單細胞植物中,所有個體或營養細胞都可能直接轉變為配子或產生配子,而在高等動物中,生殖細胞是由特殊的性腺產生的。
種類介紹
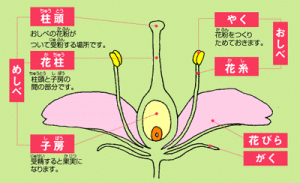 植物的有性生殖
植物的有性生殖·孤雌生殖
孤雌生殖(parthenogenesis)有些動物的生殖不需雄體參加,只是雌體產生卵。此卵不與精子結合,即可發育成新的個體,實際上這是單性生殖。其卵發育成的子代一般均為雌性個體。少數動物至今只有雌性個體,尚未發現雄性個體的存在,一生中只進行孤雌生殖,如某些輪蟲。有些動物的生殖是兩性生殖與孤雌生殖並存,如蜜蜂,其卵不受精即發育成雄蜂;卵受精則發育成雌性的工蜂和蜂王。有的動物在一生中有一段時期進行兩性生殖,另一時期進行孤雌生殖。如蚜蟲,在其生活史中,大多數時間進行孤雌生殖,只到秋末時才進行兩性生殖,產生的卵與精子結合後,休眠越冬,次年再發育成雌性蚜蟲。
·幼體生殖
幼體生殖(paedogenesis)動物的幼體生長發育成熟後,即形成成體,其生殖器官才能產生精子或卵,進行有性生殖。但某些動物的幼體雖未發育為成體,即未達性成熟,在幼體階段就可進行生殖,繁殖後代,稱為幼體生殖。幼體生殖產生的不是卵而都是幼體,因此這可以認為是胎生的一種形式。幼體生殖的母體在進行生殖時不需雄體參加,即卵不與精子結合,因此幼體生殖也可看成是孤雌生殖的一種類型。如癭蚊(Oligarcesparadoxus)除夏季進行兩性生殖外,其餘季節都進行幼體生殖。
·多胚生殖
多胚生殖(polyembryony)一個卵可產生二個或更多的胚胎,發育成多個個體的生殖方法,稱為多胚生殖。此種生殖為卵核分裂一次,以後就形成兩個胚胎,但大多數種類的卵核都經多次分裂,形成許多子核。多者可產生1600~1800個。如廣腹細蜂(Platygasterrernalis)一個卵能產生15~20個胚胎;小蜂(Litomastixtruncatellus)可產生2000個胚胎。
對比介紹
無性生殖與有性生殖的比較
·無性生殖 有性生殖——顯微鏡下的結構
有性生殖——顯微鏡下的結構1、分裂生殖,如變形蟲;2、出芽生殖,如水螅;3、孢子生殖,如根霉;4、營養生殖,如草霉。扦插、嫁接都屬於營養生殖。如:“無心插柳柳成蔭”。克隆的原意是“離體的小樹枝發育成一個植物體。”即無性繁殖的意思。高等植物的無性繁殖比較普遍,而高等動物的無性繁殖則比較困難,1997年英國科學家用克隆技術培育出了“多莉羊”,這標誌著生物新世紀的到來。
·有性生殖
由親本產生有性生殖細胞(也叫配子),經過兩性生殖細胞(如卵細胞和精子)的結合,成為合子(如受精卵),再由合子發育成為新個體的生殖方式,叫做有性生殖。有性生殖有三種:同配生殖、異配生殖和卵式生殖。而配子生殖中最高級的生殖方式是——卵式生殖。
團藻是生活在淡水中的一種能夠遊動的藻類植物。團藻的直徑只有1-2毫米。一個團藻由幾百個到幾萬個細胞構成,這些細胞都排在團藻的表面,其中只有少數的細胞具有生殖的功能。在進行有性生殖時,有的細胞長得很大,失去鞭毛,不能遊動,這種大的配子叫卵細胞;有的細胞能夠產生大量的小細胞,小細胞生有兩根鞭毛,能夠遊動,這種小配子叫做精子。兩者大小懸殊。
精子成熟時,游到卵細胞近旁,與卵細胞結合成合子(這種合子也叫做受精卵),合子外被厚壁,這種合子可以發育成新的個體——團藻。這種卵細胞與精子結合的生殖方式叫卵式生殖。
在單細胞中普遍的生殖方式是分裂生殖;生物界中普遍的生殖方式是有性生殖;高等植物和多數動物所普遍具有的一種有性生殖方式是卵式生殖。
·對比
1、無性生殖和有性生殖的根本區別是:前者不經過兩性生殖細胞的結合,由母體直接產生新個體;後者要經過兩性生殖細胞的結合,成為合子,由合子發育成新個體。
2、有性生殖與無性生殖相比具有更大的生活力和變異性。從進化的觀點看生物的生殖方式是由無性生殖向有性生殖的過渡。
生物學意義
 有性生殖——減數分裂
有性生殖——減數分裂有性生殖還能夠促進有利突變在種群中的傳播。如果一個物種有兩個個體在不同的位點上發生了有利突變,在無性生殖的種群內,這兩個突變體必將競爭,直到一個消滅為止,無法同時保留這兩個有利的突變。但在有性生殖的種群內,通過交配與重組,可以使這兩個有利的突變同時進入同一個體的基因組中,並且同時在種群中傳播。
此外進行有性生殖的物種其生活周期中都有二倍體的階段。二倍體的物種每一基因都有兩份,有一份在機能上處於備用狀態。如果這個備用的基因發生突變,成為有新的功能的基因,但此時新功能還是潛在的。通過自發的重複和有性生殖中的遺傳重組,這個新基因可與原有基因先後排列,這樣便產生一個新的基因。二倍體物種可以用這樣的方法使其基因組不斷豐富。
由於上述原因,有性生殖加速了進化的進程。在地球上生物進化的30餘億年中,前20餘億年生命停留在無性生殖階段,進化緩慢,後10億年左右進化速度明顯加快。除了地球環境的變化(例如含氧大氣的出現等)外,有性生殖的發生與發展也是一個主要的原因。現存150餘萬種生物中,從細菌到高等動植物,能進行有性生殖的種類占98%以上,就說明了這一點。
內容評價
 有性生殖
有性生殖配子是由營養個體所產生的生殖細胞,需兩兩配合後才能繼續其生活史。按配子的大小、形狀和性表現可分為3種類型:①同配生殖。配子的形態和機能完全相同,沒有性的區分。②異配生殖。有兩種類型:一種是生理的異配生殖,參加結合的配子形態上並無區別,但交配型不同,只有不同交配型的配子才能結合。另一種是形態的異配生殖,參加結合的配子形狀相同,但大小和性表現不同。③卵配生殖。相結合的雌雄配子高度特化,其大小、形態和性表現都明顯不同,成為卵和精子。卵和精子經過受精,融合為受精卵。卵配生殖是分化顯著的異配生殖。
從動、植物生殖細胞的形成過程中均有複雜的減數分裂來看,有性生殖應起源於動、植物分化前,但這還只是一種推測。
有性生殖中基因組合的廣泛變異能增加子代適應自然選擇的能力。有性生殖能促進有利突變在種群中的傳播。如果一個物種有兩個個體在不同的位點上發生了有利突變,在無性生殖的種群內,這兩個突變體必將競爭,直到一個消滅為止。但在有性生殖的種群內,通過交配與重組,可以使這兩個有利的突變同時進入同一個體的基因組中,並且同時在種群中傳播。有性生殖的物種其生活周期中都有二倍體的階段。二倍體的物種每一基因都有兩份,有一份在機能上處於備用狀態。如果這個備用的基因發生突變,通過自發的重複和有性生殖中的遺傳重組,這個新基因可與原有基因先後排列,這樣便產生一個新的基因。二倍體物種可以用這樣的方法使其基因組不斷豐富。由於上述原因,有性生殖加速了進化的進程。現存150餘萬種生物中,從細菌到高等動植物,能進行有性生殖的種類占93%以上,就說明了這一點。
