
細菌接合
正文
細菌通過細胞的暫時溝通和染色體轉移而導致基因重組的過程。細菌中導致基因重組的過程還有轉化和轉導。這兩個過程都不需要細菌細胞的直接接觸,而且基因重組的範圍更小,只限於更小的一段染色體片段和少數幾個緊密連鎖的基因。細菌接合現象是美國微生物遺傳學家J.萊德伯格和美國生物化學家兼微生物遺傳學家E.L.塔特姆於1946~1947年在大腸桿菌K-12品系中發現並證實的(見微生物遺傳學)。他們將大腸桿菌K-12品系的兩個不同的三重營養缺陷型細胞各108個混合塗布在基本培養基上,經過培養後出現少數原養型菌落。通過一系列實驗排除了回復突變、轉化和互養的可能性,從而證明這些原養型細胞是由兩個不同基因型的大腸桿菌細胞相互接觸而導致染色體DNA的轉移和重組從而產生的重組體。
英國微生物遺傳學家W.海斯和美國微生物遺傳學家萊德伯格等在1952年各自證明大腸桿菌細胞也有性別,這種性別與大腸桿菌細胞中是否存在稱為 F因子的質粒有關,這種質粒又稱為性因子或致育因子。具有F因子的細菌是染色體的供體(雄性)細菌,沒有F因子的細菌(F-)是染色體的受體(雌性)細菌。 F因子是環狀的脫氧核糖核酸(DNA)分子,大約由94500鹼基對組成,它上面存在著決定細菌細胞表面形成性傘毛的基因。當細菌接合時,供體細胞通過細胞表面的性傘毛與受體細胞相連線(圖1),與此同時供體細菌的染色體 DNA單鏈向受體細胞轉移並和受體細菌的染色體DNA發生重組(見轉化, 一般認為這種染色體DNA的轉移是通過性傘毛的孔道進行的。 單就細菌接合的生物學意義來說,它相當於高等動植物的有性生殖,但是兩者之間有以下幾點重要的區別:①高等生物中通過受精作用結合在一起的細胞一般只限於雌雄配子,它們通過減數分裂產生。細菌接合中的兩個細胞並不是通過減數分裂產生的,它們就是一般的營養細胞;②高等生物的單倍體雌雄配子通過受精作用融合成為一個合子細胞。細菌接合過程中兩個細胞只是暫時溝通而不融合;③高等動植物的合子中包含來自雌雄配子的兩套染色體,細菌接合後所形成的是部分合子,這裡面包含受體(雌性)細菌的完整的染色體和供體(雄性)細菌的染色體片段;④高等動植物的減數分裂過程中任何一個染色體的任何一個部分都有可能發生重組,細菌的部分合子中發生重組的部分只限於進入受體細菌的染色體片段;⑤高等動植物的基因重組通過染色體交換。細菌接合過程中的基因重組通過不同的方式進行,不出現聯會絲複合物和交叉(見連鎖和交換)。
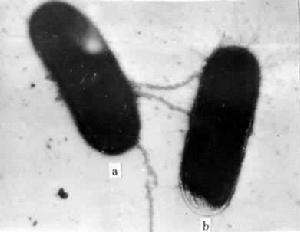 細菌接合
細菌接合關於細菌接合的知識大部分來自由法國微生物遺傳學家F.雅各布和E.沃爾曼在1956年所首創的中斷雜交實驗。所謂中斷雜交實驗就是將多標記的F-菌株的細胞和Hfr菌株的細胞相混合,培養不同時間後取少量樣品,通過劇烈攪拌使接合中的細菌細胞互相脫離,然後再在排除親本Hfr細胞的培養基上繼續培養,分析培養基上出現的菌落,看有哪些原來屬於Hfr菌株的性狀出現在F-菌株的細胞中。通過這些實驗,可以看到從混合細菌到中斷接合這一段培養時間愈長,在F-細菌中出現的Hfr菌株的性狀愈多。 一個特定的Hfr菌株的細菌和一個F-菌株的細菌接合時,首先出現在F-細胞中的供體性狀是固定不變的,並且各種性狀的出現有一定的先後次序。這表明供體染色體是從某一特定位置開始逐漸轉移的。不同的Hfr菌株的染色體轉移起點各不相同,轉移方向也不相同。
從各個菌株的各個轉移基因之間毗鄰關係相同但轉移起點和方向不同的事實可以推斷大腸桿菌的染色體是一個環狀的DNA分子,不同的Hfr菌株是由於F因子以不同方向整合到環狀染色體的不同位置上所產生的(圖2)。
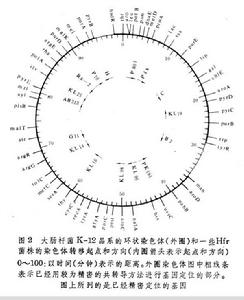 細菌接合
細菌接合在鼠傷寒沙門氏菌、綠膿桿菌、肺炎克氏桿菌、霍亂弧菌等許多細菌中也都發現有接合現象,而且像綠膿桿菌等細菌有它自己的性因子。但是到現在為止,在革蘭氏陽性細菌中還沒有發現接合現象。天藍色放線菌也能進行種內接合併且有它自己的性因子SCP1。大腸桿菌還能和沙門氏菌、奇異變形桿菌、克氏桿菌等腸道桿菌進行接合。一般親緣關係愈近的細菌重組頻率愈高,重組體也愈穩定。有些細菌本來不能進行種間的細菌接合,但經過和大腸桿菌接合而獲得 F因子後便能進行接合。在大腸桿菌中還發現另一些質粒,例如大腸桿菌素因子和某些抗藥性因子,也具有性因子的作用。
