
歷史沿革
早期發現
最早分離出DNA的弗雷德里希·米歇爾是一名瑞士醫生,他在1869年從廢棄繃帶里所殘留的膿液中,發現一些只有顯微鏡可觀察的物質。由於這些物質位於細胞核中,因此米歇爾稱之為“核素”(nuclein)。到了1919年,菲巴斯·利文進一步辨識出組成DNA的鹼基、糖類以及磷酸核苷酸單元,他認為DNA可能是許多核苷酸經由磷酸基團的聯結,而串聯在一起。不過他所提出概念中,DNA長鏈較短,且其中的鹼基是以固定順序重複排列。1937年,威廉·阿斯特伯里完成了第一張X光繞射圖,闡明了DNA結構的規律性。
1928年,弗雷德里克·格里菲斯從格里菲斯實驗中發現,平滑型的肺炎球菌,能轉變成為粗糙型的同種細菌,方法是將已死的平滑型與粗糙型活體混合在一起。這種現象稱為“轉型”。但造成此現象的因子,也就是DNA,是直到1943年,才由奧斯瓦爾德·埃弗里等人所辨識出來。1953年,阿弗雷德·赫希與瑪莎·蔡斯確認了DNA的遺傳功能,他們在赫希-蔡斯實驗中發現,DNA是T2噬菌體的遺傳物質。
組成與功能
進入20世紀時,組成蛋白質的20種胺基酸中已有12種被發現,到1940年則全部被發現。
20世紀初,德國科賽爾(1853-1927)和他的兩個學生瓊斯(1865-1935)和列文(1869-1940)的研究,弄清了核酸的基本化學結構,認為它是由許多核苷酸組成的大分子。核苷酸是由鹼基、核糖和磷酸構成的。其中鹼基有4種(腺嘌呤、鳥嘌呤、胸腺嘧啶和胞嘧啶),核糖有兩種(核糖、脫氧核糖),因此把核酸分為核糖核酸(RNA)和脫氧核糖核酸(DNA)。
列文急於發表他的研究成果,錯誤地認為4種鹼基在核酸中的量是相等的,從而推導出核酸的基本結構是由4個含不同鹼基的核苷酸連線成的四核苷酸,以此為基礎聚合成核酸,提出了“四核苷酸假說”。這個錯誤的假說,對認識複雜的核酸結構起了相當大的阻礙作用,也在一定程度上影響了人們對核酸功能的認識。
1902年,德國化學家費歇爾提出胺基酸之間以肽鏈相連線而形成蛋白質的理論,1917年他合成了由15個甘氨酸和3個亮氨酸組成的18個肽的長鏈。於是,有的科學家構想,很可能是蛋白質在遺傳中起主要作用。如果核酸參與遺傳作用,也必然是與蛋白質連在一起的核蛋白在起作用。因此,那時生物界普遍傾向於認為蛋白質是遺傳信息的載體。
到了1919年,菲巴斯·利文進一步辨識出組成DNA的鹼基、糖類以及磷酸核苷酸單元,他認為DNA可能是許多核苷酸經由磷酸基團的聯結,而串聯在一起。不過他所提出概念中,DNA長鏈較短,且其中的鹼基是以固定順序重複排列。1937年,威廉·阿斯特伯里完成了第一張X光繞射圖,闡明了DNA結構的規律性。
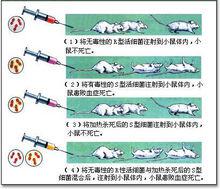 格里菲斯轉化實驗
格里菲斯轉化實驗 1928年,美國科學家弗雷德里克·格里菲斯(1877-1941)在實驗中發現,平滑型的肺炎球菌,能轉變成為粗糙型的同種細菌,方法是將已死的平滑型與粗糙型活體混合在一起。
格里菲斯用一種有莢膜、毒性強的和一種無莢膜、毒性弱的肺炎雙球菌對老鼠做實驗。他把有莢病菌用高溫殺死後與無莢的活病菌一起注入老鼠體內,結果他發現老鼠很快發病死亡,同時他從老鼠的血液中分離出了活的有莢病菌。這說明無莢菌竟從死的有莢菌中獲得了什麼物質,使無莢菌轉化為有莢菌。這種假設是否正確呢?
格里菲斯又在試管中做實驗,發現把死了的有莢菌與活的無莢菌同時放在試管中培養,無莢菌全部變成了有莢菌,並發現使無莢菌長出蛋白質莢的就是已死的有莢菌殼中遺留的核酸(因為在加熱中,莢中的核酸並沒有被破壞)。格里菲斯稱該核酸為"轉化因子"。這種現象稱為“轉化”。
但這個發現沒有得到廣泛的承認,人們懷疑當時的技術不能除淨蛋白質,殘留的蛋白質起到轉化的作用。造成此現象的因子,也就是DNA,是直到1943年,才由奧斯瓦爾德·埃弗里(O,Avery)等人所辨識出來。1953年,阿弗雷德·赫希與瑪莎·蔡斯確認了DNA的遺傳功能,他們在赫希-蔡斯實驗中發現,DNA是T噬菌體的遺傳物質。
1952年,噬菌體小組主要成員赫爾希(1908一)和他的學生蔡斯用先進的同位素標記技術,做噬菌體侵染大腸桿菌的實驗。他把大腸桿菌T2噬菌體的核酸標記上32P,蛋白質外殼標記上35S。先用標記了的T2噬菌體感染大腸桿菌,然後加以分離,結果噬菌體將帶35S標記的空殼留在大腸桿菌外面,只有噬菌體內部帶有32P標記的核酸全部注入大腸桿菌,並在大腸桿菌內成功地進行噬菌體的繁殖。這個實驗證明DNA有傳遞遺傳信息的功能,而蛋白質則是由 DNA的指令合成的。這一結果立即為學術界所接受。
美籍德國科學家德爾布呂克(1906--1981)的噬菌體小組對艾弗里的發現堅信不移。因為他們在電子顯微鏡下觀察到了噬菌體的形態和進入大腸桿菌的生長過程。噬菌體是以細菌細胞為寄主的一種病毒,個體微小,只有用電子顯微鏡才能看到它。它像一個小蝌蚪,外部是由蛋白質組成的頭膜和尾鞘,頭的內部含有DNA,尾鞘上有尾絲、基片和小鉤。當噬菌體侵染大腸桿菌時,先把尾部末端扎在細菌的細胞膜上,然後將它體內的DNA全部注入到細菌細胞中去,蛋白質空殼仍留在細菌細胞外面,再沒有起什麼作用了。進入細菌細胞後的噬菌體DNA,就利用細菌內的物質迅速合成噬菌體的DNA和蛋白質,從而複製出許多與原噬菌體大小形狀一模一樣的新噬菌體,直到細菌被徹底解體,這些噬菌體才離開死了的細菌,再去侵染其他的細菌。
幾乎與此同時,奧地利生物化學家查伽夫對核酸中的4種鹼基的含量的重新測定取得了成果。在艾弗里工作的影響下,他認為如果不同的生物種是由於DNA的不同,則DNA的結構必定十分複雜,否則難以適應生物界的多樣性。因此,他對列文的"四核苷酸假說"產生了懷疑。在1948- 1952年4年時間內,他利用了比列文時代更精確的紙層析法分離4種鹼基,用紫外線吸收光譜做定量分析,經過多次反覆實驗,終於得出了不同於列文的結果。實驗結果表明,在DNA大分子中嘌呤和嘧啶的總分子數量相等,其中腺嘌呤A與胸腺嘧啶T數量相等,鳥嘌呤G與胞嘧啶C數量相等。說明DNA分子中的鹼基A 與T、G與C是配對存在的,從而否定了“四核苷酸假說”,並為探索DNA分子結構提供了重要的線索和依據。
克里克在1957年的一場演說中,提出了分子生物學的中心法則,預測了DNA、RNA以及蛋白質之間的關係,並闡述了“轉接子假說”(即後來的tRNA)。1958年,馬修·梅瑟生與富蘭克林·史達在梅瑟生-史達實驗中,確認了DNA的複製機制。後來克里克團隊的研究顯示,遺傳密碼是由三個鹼基以不重複的方式所組成,稱為密碼子。這些密碼子所構成的遺傳密碼,最後是由哈爾·葛賓·科拉納、羅伯特·W·霍利以及馬歇爾·沃倫·尼倫伯格解出。
雙螺旋的發現
20世紀30年代後期,瑞典的科學家們就證明DNA是不對稱的。第二次世界大戰後,用電子顯微鏡測定出DNA分子的直徑約為2nm。DNA雙螺旋結構被發現後,極大地震動了學術界,啟發了人們的思想。從此,人們立即以遺傳學為中心開展了大量的分子生物學的研究。首先是圍繞著4 種鹼基怎樣排列組合進行編碼才能表達出20種胺基酸為中心開展實驗研究。
20世紀50年代,DNA雙螺旋結構被闡明,揭開了生命科學的新篇章,開創了科學技術的新時代。隨後,遺傳的分子機理――DNA複製、遺傳密碼、遺傳信息傳遞的中心法則、作為遺傳的基本單位和細胞工程藍圖的基因以及基因表達的調控相繼被認識。至此,人們已完全認識到掌握所有生物命運的東西就是DNA和它所包含的基因,生物的進化過程和生命過程的不同,就是因為DNA和基因運作軌跡不同所致。
1953年4月25日,英國的《自然》雜誌刊登了美國的沃森和英國的克里克在英國劍橋大學合作的研究成果:DNA雙螺旋結構的分子模型,這一成果後來被譽為20世紀以來生物學方面最偉大的發現,標誌著分子生物學的誕生。
 沃森
沃森 沃森(1928一)在中學時代是一個極其聰明的孩子,15歲時便進入芝加哥大學學習。當時,由於一個允許較早人學的實驗性教育計畫,使沃森有機會從各個方面完整地攻讀生物科學課程。
在大學期間,沃森在遺傳學方面雖然很少有正規的訓練,但自從閱讀了薛丁格的《生命是什麼?--活細胞的物理面貌》這本進化論的理論基礎書籍,促使他去“發現基因的秘密”。他善於集思廣益,博取眾長,善於用他人的思想來充實自己。只要有便利的條件,不必強迫自己學習整個新領域,也能得到所需要的知識。
沃森22歲取得博士學位,然後被送往歐洲攻讀博士後研究員。為了完全搞清楚一個病毒基因的化學結構,他到丹麥哥本哈根實驗室學習化學。有一次他與導師一起到義大利那不勒斯參加一次生物大分子會議,有機會聽英國物理生物學家莫里斯·威爾金斯(1916--2004)的演講,看到了威爾金斯的DNAX射線衍射照片。從此,尋找解開DNA結構的鑰匙的念頭在沃森的頭腦中索回。什麼地方可以學習分析X射線衍射圖呢?於是他又到英國劍橋大學卡文迪什實驗室學習,在此期間沃森認識了克里克。
 克里克
克里克 克里克(1916-2004)上中學時對科學充滿熱情,1937年畢業於倫敦大學。1946年,他閱讀了埃爾溫·薛丁格《生命是什麼?-活細胞的物理面貌》一書,決心把物理學知識用於生物學的研究,從此對生物學產生了興趣。1947年他重新開始了研究生的學習,1949年他同佩魯茲一起使用X射線技術研究蛋白質分子結構,於是在此與沃森相遇了。
當時克里克比沃森大12歲,還沒有取得博士學位。但他們談得很投機,沃森感到在這裡居然能找到一位懂得DNA比蛋白質更重要的人,真是三生有幸。同時沃森感到在他所接觸的人當中,克里克是最聰明的一個。他們每天交談至少幾個小時,討論學術問題。兩個人互相補充,互相批評以及相互激發出對方的靈感。
他們認為解決DNA分子結構是打開遺傳之謎的關鍵。只有藉助於精確的X射線衍射資料,才能更快地弄清DNA的結構。為了搞到DNAX射線衍射資料,克里克請威爾金斯到劍橋來度周末。在交談中威爾金斯接受了DNA結構是螺旋型的觀點,還談到他的合作者富蘭克林(1920--1958,女)以及實驗室的科學家們,也在苦苦思索著DNA結構模型的問題。從1951年11月至1953年4月的18個月中,沃森、克里克同威爾金斯、富蘭克林之間有過幾次重要的學術交往。
1951年11月,沃森聽了富蘭克林關於DNA結構的較詳細的報告後,深受啟發,具有一定晶體結構分析知識的沃森和克里克認識到,要想很快建立 DNA結構模型,只能利用別人的分析數據。他們很快就提出了一個三股螺旋的DNA結構的構想。1951年底,他們請威爾金斯和富蘭克林來討論這個模型時,富蘭克林指出他們把DNA的含水量少算了一半,於是第一次設立的模型宣告失敗。
有一天,沃森又到國王學院威爾金斯實驗室,立刻興奮起來、心跳也加快了,因為這種圖像比以前得到的“A型”簡單得多,只要稍稍看一下“B型”的X射線衍射照片,再經簡單計算,就能確定DNA分子內多核苷酸鏈的數目了。
克里克請數學家幫助計算,結果表明嘌呤有吸引嘧啶的趨勢。他們根據這一結果和從查伽夫處得到的核酸的兩個嘌呤和兩個嘧啶兩兩相等的結果,形成了鹼基配對的概念。
他們苦苦地思索4種鹼基的排列順序,一次又一次地在紙上畫鹼基結構式,擺弄模型,一次次地提出假設,又一次次地推翻自己的假設。
有一次,沃森又在按著自己的構想擺弄模型,他把鹼基移來移去尋找各種配對的可能性。突然,他發現由兩個氫鍵連線的腺嘌呤一胸腺嘧啶對竟然和由3個氫鍵連線的鳥嘌呤一胞嘧啶對有著相同的形狀,於是精神為之大振。因為嘌呤的數目為什麼和嘧啶數目完全相同這個謎就要被解開了。查伽夫規律也就一下子成了 DNA雙螺旋結構的必然結果。因此,一條鏈如何作為模板合成另一條互補鹼基順序的鏈也就不難想像了。那么,兩條鏈的骨架一定是方向相反的。
經過沃森和克里克緊張連續的工作,很快就完成了DNA金屬模型的組裝。從這模型中看到,DNA由兩條核苷酸鏈組成,它們沿著中心軸以相反方向相互纏繞在一起,很像一座螺旋形的樓梯,兩側扶手是兩條多核苷酸鏈的糖一磷基因交替結合的骨架,而踏板就是鹼基對。由於缺乏準確的X射線資料,他們還不敢斷定模型是完全正確的。
下一步的科學方法就是把根據這個模型預測出的衍射圖與X射線的實驗數據作一番認真的比較。他們又一次打電話請來了威爾金斯。不到兩天工夫,威爾金斯和富蘭克林就用X射線數據分析證實了雙螺旋結構模型是正確的,並寫了兩篇實驗報告同時發表在英國《自然》雜誌上。1962年,沃森、克里克和威爾金斯獲得了諾貝爾醫學和生理學獎,而富蘭克林因患癌症於1958年病逝而未被授予該獎。
基因工程
1967年,遺傳密碼全部被破解,基因從而在DNA分子水平上得到新的概念。它表明:基因實際上就是DNA大分子中的一個片段,是控制生物性狀的遺傳物質的功能單位和結構單位。在這個單位片段上的許多核苷酸不是任意排列的,而是以有含意的密碼順序排列的。一定結構的DNA,可以控制合成相應結構的蛋白質。蛋白質是組成生物體的重要成分,生物體的性狀主要是通過蛋白質來體現的。因此,基因對性狀的控制是通過DNA控制蛋白質的合成來實現的。在此基礎上相繼產生了基因工程、酶工程、發酵工程、蛋白質工程等技術。
1972年,美國科學家保羅.伯格首次成功地重組了世界上第一批DNA分子,標誌著DNA重組技術――基因工程作為現代生物工程的基礎,成為現代生物技術和生命科學的基礎與核心。
到了20世紀70年代中後期,由於出現了工程菌以及實現DNA重組和後處理都有工程化的性質,基因工程或遺傳工程作為DNA重組技術的代名詞被廣泛使用。
到20世紀末,DNA重組技術最大的套用領域在醫藥方面,包括活性多肽、蛋白質和疫苗的生產,疾病發生機理、診斷和治療,新基因的分離以及環境監測與淨化。
理化性質
 第一張DNA照片
第一張DNA照片 DNA是一種長鏈聚合物,組成單位為四種脫氧核苷酸,即:
腺嘌呤脫氧核苷酸(dAMP )、胸腺嘧啶脫氧核苷酸(dTMP )、胞嘧啶脫氧核苷酸(dCMP )、鳥嘌呤脫氧核苷酸(dGMP )。
脫氧核糖核酸是一種由核苷酸重複排列組成的長鏈聚合物,寬度約22到24埃(2.2到2.4納米),每一個核苷酸單位則大約長3.3埃(0.33納米)。在整個脫氧核糖核酸聚合物中,可能含有數百萬個相連的核苷酸。例如人類細胞中最大的1號染色體中,就有2億2千萬個鹼基對。通常在生物體內,脫氧核糖核酸並非單一分子,而是形成兩條互相配對並緊密結合,且如藤蔓般地纏繞成雙螺旋結構的分子。每個核苷酸分子的其中一部分會相互連結,組成長鏈骨架;另一部分稱為鹼基,可使成對的兩條脫氧核糖核酸相互結合。所謂核苷酸,是指一個核苷加上一個或多個磷酸基團,核苷則是指一個鹼基加上一個糖類分子。
脫氧核糖核酸骨架是由磷酸與糖類基團互動排列而成。組成脫氧核糖核酸的糖類分子為環狀的2-脫氧核糖,屬於五碳糖的一種。磷酸基團上的兩個氧原子分別接在五碳糖的3號及5號碳原子上,形成磷酸雙酯鍵。這種兩側不對稱的共價鍵位置,使每一條脫氧核糖核酸長鏈皆具方向性。雙螺旋中的兩股核苷酸互以相反方向排列,這種排列方式稱為反平行。脫氧核糖核酸鏈上互不對稱的兩末端一邊叫做5'端,另一邊則稱3'端。脫氧核糖核酸與RNA最主要的差異之一,在於組成糖分子的不同,DNA為2-脫氧核糖,RNA則為核糖。
而脫氧核糖(五碳糖)與磷酸分子藉由酯鍵相連,組成其長鏈骨架,排列在外側,四種鹼基排列在內側。每個糖分子都與四種鹼基里的其中一種相連,這些鹼基沿著DNA長鏈所排列而成的序列,可組成遺傳密碼,指導蛋白質的合成。讀取密碼的過程稱為轉錄,是以DNA雙鏈中的一條單鏈為模板轉錄出一段稱為mRNA(信使RNA)的核酸分子。多數RNA帶有合成蛋白質的訊息,另有一些本身就擁有特殊功能,例如rRNA、snRNA與siRNA。
在細胞內,DNA能與蛋白質結合形成染色體,整組染色體則統稱為染色體組。對於人類而言,正常的人體細胞中含有46條染色體。染色體在細胞分裂之前會先在分裂間期完成複製,細胞分裂間期又可劃分為:G1期-DNA合成前期、S期-DNA合成期、G2-DNA合成後期。對於真核生物,如動物、植物及真菌而言,染色體主要存在於細胞核內;而對於原核生物,如細菌而言,則主要存在於細胞質中的擬核內。染色體上的染色質蛋白,如組織蛋白,能夠將DNA進行組織並壓縮,以幫助DNA與其他蛋白質進行互動作用,進而調節基因的轉錄。
DNA是高分子聚合物,DNA溶液為高分子溶液,具有很高的粘度,可被甲基綠染成綠色。DNA對紫外線(260nm)有吸收作用,利用這一特性,可以對DNA進行含量測定。當核酸變性時,吸光度升高,稱為增色效應;當變性核酸重新復性時,吸光度又會恢復到原來的水平。較高溫度、有機溶劑、酸鹼試劑、尿素、醯胺等都可以引起DNA分子變性,即DNA雙鏈鹼基間的氫鍵斷裂,雙螺旋結構解開—也稱為DNA的解螺旋。
分子結構
DNA是由許多脫氧核苷酸按一定鹼基順序彼此用3’,5’-磷酸二酯鍵相連構成的長鏈。大多數DNA含有兩條這樣的長鏈,也有的DNA為單鏈,如大腸桿菌噬菌體φX174、G4、M13等。DNA有環形DNA和鏈狀DNA之分。在某些類型的DNA中,5-甲基胞嘧啶可在一定限度內取代胞嘧啶,其中小麥胚DNA的5-甲基胞嘧啶特別豐富。在某些噬菌體中,5-羥甲基胞嘧啶取代了胞嘧啶。40年代後期,查伽夫(E.Chargaff)發現不同物種DNA的鹼基組成不同,但其中的腺嘌呤數等於其胸腺嘧啶數(A=T),鳥嘌呤數等於胞嘧啶數(G=C),因而嘌呤數之和等於嘧啶數之和,一般用幾個層次描繪DNA的結構。
一級結構
 一級結構
一級結構 是指構成核酸的四種基本組成單位——脫氧核糖核苷酸(核苷酸),通過3',5'-磷酸二酯鍵彼此連線起來的線形多聚體,以及其基本單位-脫氧核糖核苷酸的排列順序。每一種脫氧核糖核苷酸由三個部分所組成:一分子含氮鹼基+一分子五碳糖(脫氧核糖)+一分子磷酸根。核酸的含氮鹼基又可分為四類:腺嘌呤(adenine,縮寫為A),胸腺嘧啶(thymine,縮寫為T),胞嘧啶(cytosine,縮寫為C)和鳥嘌呤(guanine,縮寫為G)。DNA的四種含氮鹼基組成具有物種特異性。即四種含氮鹼基的比例在同物種不同個體間是一致的,但在不同物種間則有差異。DNA的四種含氮鹼基比例具有奇特的規律性,每一種生物體DNA中 A=T ,C=G 查伽夫規則(即 鹼基互補配對原則)。
二級結構
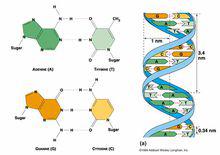 二級結構
二級結構 是指兩條脫氧多核苷酸鏈反向平行盤繞所形成的雙螺旋結構。DNA的二級結構分為兩大類:一類是右手螺旋,如A-DNA、B-DNA、C-DNA、D-DNA等;另一類是左手雙螺旋,如Z-DNA。詹姆斯·沃森與佛朗西斯·克里克所發現的雙螺旋,是稱為B型的水結合型DNA,在細胞中最為常見(如圖)。也有的DNA為單鏈,一般見於病毒,如大腸桿菌噬菌體φX174、G4、M13等。有的DNA為環形,有的DNA為線形。在鹼A與T之間可以形成兩個氫鍵,G與C之間可以形成三個氫鍵,使兩條多聚脫氧核苷酸形 成互補的雙鏈,由於組成鹼基對的兩個鹼基的分布不在一個平面上,氫鍵使鹼基對沿長軸旋轉一定角度,使鹼基的形狀像螺旋槳葉片的樣子,整個DNA分子形成雙螺旋纏繞狀。鹼基對之間的距離是0.34nm,10個鹼基對轉一周,故旋轉一周(螺距)是3.4nm,這是β-DNA的結構,在生物體內自然生成的DNA幾乎都是以β-DNA結構存在。
三級結構
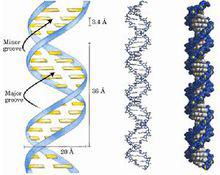 三級結構
三級結構 是指DNA中單鏈與雙鏈、雙鏈之間的相互作用形成的三鏈或四鏈結構。如H-DNA或R-環等三級結構。DNA的三級結構是指DNA進一步扭曲盤繞所形成的特定空間結構,也稱為超螺旋結構。DNA的超螺旋結構可分為正、負超螺旋兩大類,並可互相轉變。超螺旋是克服張力而形成的。當DNA雙螺旋分子在溶液中以一定構象自由存在時,雙螺旋處於能量最低狀態此為鬆弛態。如果使這種正常的DNA分子額外地多轉幾圈或少轉幾圈,就是雙螺旋產生張力,如果DNA分子兩端是開放的,這種張力可通過鏈的轉動而釋放出來,DNA就恢復到正常的雙螺旋狀態。但如果DNA分子兩端是固定的,或者是環狀分子,這種張力就不能通過鏈的旋轉釋放掉,只能使DNA分子本身發生扭曲,以此抵消張力,這就形成超螺旋,是雙螺旋的螺旋。
四級結構
核酸以反式作用存在(如核糖體、剪接體),這可看作是核酸的 四級水平的結構。
拓撲結構
也是DNA存在的一種形式。DNA的拓撲結構是指在DNA雙螺旋的基礎上,進一步扭曲所形成的特定空間結構。超螺旋結構是拓撲結構的主要形式,它可以分為正超螺旋和負超螺旋兩類,在相應條件下,它們可以相互轉變。
其他結構
原核細胞的遺傳物質是一個長DNA分子,但是原核細胞沒有真正的細胞核。真核細胞核中有不止一個染色體,每個染色體也只含一個DNA分子。不過它們一般都比原核細胞中的DNA分子大而且和蛋白質結合在一起。DNA分子的功能是貯存決定物種的所有蛋白質和RNA結構的全部遺傳信息;策劃生物有次序地合成細胞和組織組分的時間和空間;確定生物生命周期自始至終的活性和確定生物的個性。除染色體DNA外,有極少量結構不同的DNA存在於真核細胞的線粒體和葉綠體中。DNA病毒的遺傳物質也是DNA。
主要類別
單鏈DNA
單鏈DNA(single-stranded DNA)大部分DNA以雙螺旋結構存在,但一經熱或鹼處理就會變為單鏈狀態。單鏈DNA就是指以這種狀態存在的DNA。單鏈DNA在分子流體力學性質、吸收光譜、鹼基反應性質等方面都和雙鏈DNA不同。某些噬菌體粒子內含有單鏈環狀的DNA,這樣的噬菌體DNA在細胞內增殖時則形成雙鏈DNA。
閉環DNA
閉環DNA(closed circular DNA)沒有斷口的雙鏈環狀DNA,亦稱為超螺旋DNA。由於具有螺旋結構的雙鏈各自閉合,結果使整個DNA分子進一步旋曲而形成三級結構。另外如果一條或二條鏈的不同部位上產生一個斷口,就會成為無旋曲的開環DNA分子。從細胞中提取出來的質粒或病毒DNA都含有閉環和開環這二種分子。可根據兩者與色素結合能力的不同,而將兩者分離開來。
連線DNA
連線DNA (Linker DNA):核小體中除147bp核心DNA 外的所有DNA。
模板DNA
模板DNA可以是單鏈分子,也可以是雙鏈分子,可以是線狀分子,也可以是環狀分子(線狀分子比環狀分子的擴增效果稍好)。就模板DNA而言,影響PCR的主要因素是模板的數量和純度。
互補DNA
互補DNA(cDNA,complementary DNA)構成基因的雙鏈DNA分子用一條單鏈作為模板,轉錄產生與其序列互補的信使RNA分子,然後在反轉錄酶的作用下,以mRNA分子為模板,合成一條與mRNA序列互補的單鏈DNA,最後再以單鏈DNA為模板合成另一條與其互補的單鏈DNA,兩條互補的單鏈DNA分子組成一個雙鏈cDNA分子.因此,雙鏈cDNA分子的序列同轉錄產生的mRNA分子的基因是相同的.所以一個cDNA分子就代表一個基因.但是cDNA仍不同於基因,因為基因在轉錄產生mRNA時,一些不編碼的序列即內含子被刪除了,保留的只是編碼序列,即外顯子.所以cDNA序列都比基因序列要短得多,因為cDNA中不包括基因的非編碼序列---內含子。
技術發展
垃圾DNA
一項針對基因組進行的廣泛比較研究顯示,問題的答案可能就隱藏在生物的垃圾脫氧核糖核酸(DNA)中。美國科學家發現,生物越複雜,其攜帶的垃圾DNA就越多,而恰恰是這些沒有編碼的“無用”DNA幫助高等生物進化出了複雜的機體。
自從第一個真核生物——包括從酵母到人類的有細胞核的生物——的基因組被破譯以來,科學家一直想知道,為什麼生物的大多數DNA並沒有形成有用的基因。從突變保護到染色體的結構支撐,對於這種所謂的垃圾DNA的可能解釋有許多種。但是2004年從人類、小鼠和大鼠身上得到的完全一致的關於垃圾DNA的研究結果卻表明,在這一區域中可能包含有重要的調節機制,從而能夠控制基礎的生物化學反應和發育進程,這將幫助生物進化出更為複雜的機體。與簡單的真核生物相比,複雜生物有更多的基因不會發生突變的事實無疑極大地強化了這一發現。
為了對這一問題有更深的了解,由美國加利福尼亞大學聖塔克魯斯分校(UCSC)的計算生物學家David Haussler領導的一個研究小組,對5種脊椎動物——人、小鼠、大鼠、雞和河豚——的垃圾DNA序列與4種昆蟲、兩種蠕蟲和7種酵母的垃圾DNA序列進行了比較。研究人員從對比結果中得到了一個驚人的模式:生物越複雜,垃圾DNA似乎就越重要。
這其中暗含的可能性在於,如果不同種類的生物具有相同的DNA,那么這些DNA必定是用來解決一些關鍵性的問題的。酵母與脊椎動物共享了一定數量的DNA,畢竟它們都需要製造蛋白質,但是只有15%的共有DNA與基因無關。研究小組在2005年7月14日的《基因組研究》雜誌網路版上報告說,他們將酵母與更為複雜的蠕蟲進行了比較,後者是一種多細胞生物,發現有40%的共有DNA沒有被編碼。隨後,研究人員又將脊椎動物與昆蟲進行了對比,這些生物比蠕蟲更為複雜,結果發現,有超過66%的共有DNA包含有沒有編碼的DNA。
參與該項研究工作的UCSC計算生物學家Adam Siepel指出,有關蠕蟲的研究結果需要慎重對待,這是由於科學家僅僅對其中的兩個基因組進行了分析。儘管如此,Siepel還是認為,這一發現有力地支持了這樣一種理論,即脊椎動物和昆蟲的生物複雜性的增加主要是由於基因調節的精細模式。
DNA探針
DNA探針是最常用的核酸探針,指長度在幾百鹼基對以上的雙鏈DNA或單鏈DNA探針。現已獲得DNA探針數量很多,有細菌、病毒、原蟲、真菌、動物和人類細胞DNA探針。這類探針多為某一基因的全部或部分序列,或某一非編碼序列。這些DNA片段須是特異的,如細菌的毒力因子基因探針和人類Alu探針。這些DNA探針的獲得有賴於分子克隆技術的發展和套用。以細菌為例,加之分子雜交技術的高敏感性,分子雜交在臨床微生物診斷上具有廣闊的前景。
細菌的基因組大小約5×106bp,約含3000個基因。各種細菌之間絕大部分DNA是相同的,要獲得某細菌特異的核酸探針,通常要採取建立細菌基因組DNA文庫的辦法,即將細菌DNA切成小片段後分別克隆得到包含基因組的全信息的克隆庫。然後用多種其它菌種的DNA作探針來篩選,產生雜交信號的克隆被剔除,最後剩下的不與任何其它細菌雜交的克隆則可能含有該細菌特異性DNA片段。將此重組質粒標記後作探針進一步鑑定,亦可經DNA序列分析鑑定其基因來源和功能。因此要得到一種特異性DNA探針,常常是比較繁瑣的。探針DNA克隆的篩選也可採用血清學方法,所不同的是所建DNA文庫為可表達性,克隆菌落或噬斑經裂解後釋放出表達抗原,然後用來源細菌的多克隆抗血清篩選陽性克隆,所得到多個陽性克隆再經其它細菌的抗血清篩選,最後只與本細菌抗血清反應的表達克隆即含有此細菌的特異性基因片段,它所編碼的蛋白是該菌種所特有的。用這種表達文庫篩選得到的顯然只是特定基因探針。
DNA修復
DNA修復(DNA repairing)是細胞對DNA受損傷後的一種反應,這種反應可能使DNA結構恢復原樣,重新能執行它原來的功能;但有時並非能完全消除DNA的損傷,只是使細胞能夠耐受這DNA的損傷而能繼續生存。也許這未能完全修復而存留下來的損傷會在適合的條件下顯示出來(如細胞的癌變等),但如果細胞不具備這修復功能,就無法對付經常在發生的DNA損傷事件,就不能生存。所以研究DNA修復也是探索生命的一個重要方面,而且與軍事醫學、腫瘤學等密切相關。對不同的DNA損傷,細胞可以有不同的修復反應。
DNA複製
DNA複製是指DNA雙鏈在細胞分裂以前進行的複製過程,複製的結果是一條雙鏈變成兩條一樣的雙鏈(如果複製過程正常的話),每條雙鏈都與原來的雙鏈一樣。這個過程是通過名為半保留複製的機制來得以順利完成的。複製可以分為以下幾個階段:
 電子顯微鏡下的DNA
電子顯微鏡下的DNA 起始階段:解旋酶在局部展開雙螺旋結構的DNA分子為單鏈,引物酶辨認起始位點,以解開的一段DNA為模板,按照5'到3'方向合成RNA短鏈。形成RNA引物。
DNA片段的生成:在引物提供了3'-OH末端的基礎上,DNA聚合酶催化DNA的兩條鏈同時進行複製過程,由於複製過程只能由5'->3'方向合成,因此一條鏈能夠連續合成,另一條鏈分段合成,其中每一段短鏈成為岡崎片段(Okazaki fragments)。
RNA引物的水解:當DNA合成一定長度後,DNA聚合酶水解RNA引物,補填缺口。
DNA連線酶將DNA片段連線起來,形成完整的DNA分子。
最後DNA新合成的片段在旋轉酶的幫助下重新形成螺旋狀。
PCR複製技術
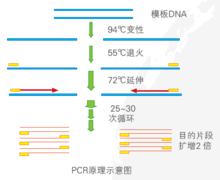 PCR原理示意圖
PCR原理示意圖 在雙螺旋的DNA中,分子鏈是由互補的核苷酸配對組成的,兩條鏈依靠氫鏈結合在一起。由於氫鏈鏈數的限制,DNA的鹼基排列配對方式只能是A對T(由兩個氫鍵相連)或C對G(由三個氫鏈相連)。因此,一條鏈的鹼基序列就可以決定了另一條的鹼基序列,因為每一條鏈的鹼基對和另一條鏈的鹼基對都必須是互補的。在DNA複製時也是採用這種互補配對的原則進行的:當DNA雙螺旋被展開時,每一條鏈都用作一個模板,通過互補的原則補齊另外的一條鏈,即半保留複製。
DNA重組
重組DNA是一種人工合成的脫氧核糖核酸。它是把一般不同時出現的DNA序列組合到一起而產生的。從遺傳工程的觀點來看重組DNA是把相關的DNA添加到已有生物的基因組中,比如細菌的質粒中,其目的是為了改變或者添加特別是的特性,比如免疫。重組DNA與遺傳重組不是一回事。它不是重組細胞內或者染色體上已經存在的基因組,而完全是通過外部工程達到的。重組蛋白質是從重組DNA合成出來的蛋白質。
重組DNA技術是1973年由斯坦利·諾曼·科恩和赫伯特·玻意爾設計的。1974年他們發表了他們的設計。在這篇論文中他們描述了分離和放大基因或者DNA片段,然後精確地把它們插入其它細胞中,由此製造出轉基因細菌。沃納·亞伯、丹尼爾·那森斯和漢彌爾頓·史密斯發明了限制酶才使得重組DNA技術可行,為此他們獲得了1978年諾貝爾醫學獎。
超速離心
近代質粒DNA分離純化以從大腸桿菌中分離為代表,鑒於大腸桿菌( E.coli)在分子生物學研究中的重要地位,從大腸桿菌( E.coli)中分離純化質粒DNA(Plasmid DNA)成為超離心技術中一個重要課題。而質粒DNA的快速分離純化又對超離心設備(超速離心機、轉頭和附屬設備)提出了更高要求。
針對 E.coli的顯微結構待點,在進行超離心分離純化質粒DNA之前的預處理順序是:
沉澱物可以在加入TE緩衝液(10mM Tris-HCL, lmM EDTA,pH8.0)後分子篩技術去除蛋白和RNA; 也可以用超速離心法去除蛋白質和RNA,去級狀DNA或DNA斷片。
質粒DNA超速離心的分離方法
傳統的分離方法:數年前,由於受設備條件限制,質粒DNA的分離一般用CsCl平衡等密度離心法,自形成梯度。以10~12ml單管容量為例,用甩平轉頭分離,36.000rpm×60小時,用角式轉頭分離45,000rpm×36小時,前者包括加減速在內共用去1.3億轉驅動部壽命,後者也要用去1億轉驅動部壽命,這對當時超速離心機總壽命為100~200億轉來看,無疑每次實驗費用過高,加上CsCl用量多、價格貴等因素,使這類分離純化工作成為非常昂貴的實驗。
質粒DNA超速離心分離的最新進展
1、超速垂直管轉頭的離心分離(欽合金或碳纖維製造的):從1975年垂直管轉頭向世後,最高轉速從50,000rpm到120,000rpm,RCFmax可達700,000Xg,90年代開發的新機型和轉頭己能夠使質粒DNA垂直管離心分離實驗做起來得心應手。
2、近垂直管轉頭離心分離:為了消除垂直管轉頭用於質粒DNA離心在壁部形成的RNA沉澱對已形成的DNA區帶的污染,同時也為了改進一般斜角式轉頭(傾角25——35)由於沉降距離較長,因而分離時間也較長的缺點,近幾年開發了多種近垂直管轉頭(即Near VerticalTube Rot時,簡稱NVT轉頭或Neo Angle Rotor,小假角轉頭,簡稱NT).它們的離心管縱剖面中心軸線與離心機驅動軸線之間夾角在7.5——10之間,轉速從65,000rpm到120,OOOrpm,RCFmax可達646,000×g單管容量從2ml至13.5ml。NVT(或NT)轉頭的開發主要是為質粒DNA分離而設計,當然它也適用於線粒體DNA、染色體DNA、RNA及血清脂蛋白的分離·純化。
3、不連續階梯梯度分離:質校DNA分離純化傳統方法是採用金管CsCl自形成梯度平衡等密度離心法,離心開始時金管CsCl密度均一,樣品均勻分布其中。
互動作用
脫氧核糖核酸若要發揮其功用,必須依賴與蛋白質之間的互動作用,有些蛋白質的作用不具專一性,有些則只專門與個別的脫氧核糖核酸序列結合。聚合酶在各類酵素中尤其重要,此種蛋白質可與脫氧核糖核酸結合,並作用於轉錄或脫氧核糖核酸複製過程。
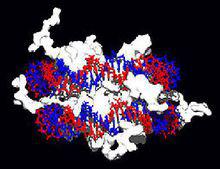 脫氧核糖核酸
脫氧核糖核酸 脫氧核糖核酸與組織蛋白(右圖白色部分)的互動作用,這種蛋白質中的鹼性胺基酸(左下藍色),可與脫氧核糖核酸上的酸性磷酸基團結合(右下紅色)。
結構蛋白可與脫氧核糖核酸結合,是非專一性脫氧核糖核酸-蛋白質互動作用的常見例子。染色體中的結構蛋白與脫氧核糖核酸組合成複合物,使脫氧核糖核酸組織成緊密結實的染色質構造。對真核生物來說,染色質是由脫氧核糖核酸與一種稱為組織蛋白的小型鹼性蛋白質所組合而成;而原核生物體內的此種結構,則摻雜了多種類型的蛋白質。
雙股脫氧核糖核酸可在組織蛋白的表面上附著並纏繞整整兩圈,以形成一種稱為核小體的盤狀複合物。組織蛋白里的鹼性殘基,與脫氧核糖核酸上的酸性糖磷酸骨架之間可形成離子鍵,使兩者發生非專一性互動作用,也使複合物中的鹼基序列相互分離。
在鹼性胺基酸殘基上所發生的化學修飾有甲基化、磷酸化與乙醯化等,這些化學作用可使脫氧核糖核酸與組織蛋白之間的作用強度發生變化,進而使脫氧核糖核酸與轉錄因子接觸的難易度改變,影響轉錄作用的速率。其他位於染色體內的非專一性脫氧核糖核酸結合蛋白,還包括一種能優先與脫氧核糖核酸結合,並使其扭曲的高移動性群蛋白。這類蛋白質可以改變核小體的排列方式,產生更複雜的染色質結構。
脫氧核糖核酸結合蛋白中有一種專門與單股脫氧核糖核酸結合的類型,稱為單股脫氧核糖核酸結合蛋白。人類的複製蛋白A是此類蛋白中獲得較多研究的成員,作用於多數與解開雙螺旋有關的過程,包括脫氧核糖核酸複製、重組以及脫氧核糖核酸修復。這類結合蛋白可固定單股脫氧核糖核酸,使其變得較為穩定,以避免形成莖環(stem-loop),或是因為核酸酶的作用而水解。
相對而言,其他的蛋白質則只能與特定的脫氧核糖核酸序列進行專一性結合。大多數關於此類蛋白質的研究集中於各種可調控轉錄作用的轉錄因子。這類蛋白質中的每一種,都能與特定的脫氧核糖核酸序列結合,進而活化或抑制位於啟動子附近序列的基因轉錄作用。轉錄因子有兩種作用方式,第一種可以直接或經由其他中介蛋白質的作用,而與負責轉錄的RNA聚合酶結合,再使聚合酶與啟動子結合,並開啟轉錄作用。第二種則與專門修飾組織蛋白的酵素結合於啟動子上,使脫氧核糖核酸模板與聚合酶發生接觸的難度改變。
由於目標脫氧核糖核酸可能散布在生物體中的整個基因組中,因此改變一種轉錄因子的活性可能會影響許多基因的運作。這些轉錄因子也因此經常成為信號傳遞過程中的作用目標,也就是作為細胞反映環境改變,或是進行分化和發育時的媒介。具專一性的轉錄因子會與脫氧核糖核酸發生互動作用,使脫氧核糖核酸鹼基的周圍產生許多接觸點,讓其他蛋白質得以“讀取”這些脫氧核糖核酸序列。多數的鹼基互動作用發生在大凹槽,也就是最容易從外界接觸鹼基的部位。
濺射法製備 a-GaN 薄膜的光學性質也類似於脫氧核糖核酸屬化學成分。
套用領域
身份鑑定
鑑定親子關係用得最多的是DNA分型鑑定。人的血液、毛髮、唾液、口腔細胞等都可以用於用親子鑑定,十分方便。
一個人有23對(46條)染色體,同一對染色體同一位置上的一對基因稱為等位基因,一般一個來自父親,一個來自母親。如果檢測到某個DNA位點的等位基因,一個與母親相同,另一個就應與父親相同,否則就存在疑問了。
利用DNA進行親子鑑定,只要作十幾至幾十個DNA位點作檢測,如果全部一樣,就可以確定親子關係,如果有3個以上的位點不同,則可排除親子關係,有一兩個位點不同,則應考慮基因突變的可能,加做一些位點的檢測進行辨別。DNA親子鑑定,否定親子關係的準確率幾近100%,肯定親子關係的準確率可達到99.99%。
DNA(脫氧核糖核酸)是人身體內細胞的原子物質。每個原子有46個染色體,另外,男性的精子細胞和女性的卵子,各有23個染色體,當精子和卵子結合的時候。這46個原子染色體就製造一個生命,因此,每人從生父處繼承一半的分子物質,而另一半則從生母處獲得。
DNA親子鑑定測試與傳統的血液測試有很大的不同。它可以在不同的樣本上進行測試,包括血液,腮腔細胞,組織細胞樣本和精液樣本。由於血液型號,例如A型,B型,O型或RH型,在人口中比較普遍,用於分辨每一個人,便不如DNA親子鑑定測試有效。除了真正雙胞胎外,每人的DNA是獨一無二的.。由於它是這樣獨特,就好像指紋一樣,用於親子鑑定,DNA是最為有效的方法。我們的結果通常是比法庭上要求的還準確10到100倍。
通過遺傳標記的檢驗與分析來判斷父母與子女是否親生關係,稱之為親子試驗或親子鑑定。DNA是人體遺傳的基本載體,人類的染色體是由DNA構成的,每個人體細胞有23對(46條)成對的染色體,其分別來自父親和母親。夫妻之間各自提供的23條染色體,在受精後相互配對,構成了23對(46條)孩子的染色體。如此循環往復構成生命的延續。
由於人體約有30億個鹼基對構成整個染色體系統,而且在生殖細胞形成前的互換和組合是隨機的,所以世界上沒有任何兩個人具有完全相同的30億個核苷酸的組成序列,這就是人的遺傳多態性。儘管遺傳多態性的存在,但每一個人的染色體必然也只能來自其父母,這就是DNA親子鑑定的理論基礎。
傳統的血清方法能檢測紅細胞血型、白細胞血型、血清型和紅細胞酶型等,這些遺傳學標誌為蛋白質(包括糖蛋白)或多肽,容易失活而導致檢材得不到理想的檢驗結果。此外,這些遺傳標誌均為基因編碼的產物,多態信息含量(PIC)有限,不能反映DNA編碼區的多態性,且這些遺傳標誌存在生理性、病理性變異(如A型、O型血的人受大腸桿菌感染後,B抗原可能呈陽性。因此,其套用價值有限。
DNA檢驗可彌補血清學方法的不足,故受到了法醫物證學工作者的高度關注,近幾年來,人類基因組研究的進展日新月異,而分子生物學技術也不斷完善,隨著基因組研究向各學科的不斷滲透,這些學科的進展達到了前所未有的高度。在法醫學上,STR位點和單核苷酸(SNP)位點檢測分別是第二代、第三代DNA分析技術的核心,是繼RFLPs(限制性片段長度多態性)VNTRs(可變數量串聯重複序列多態性)研究而發展起來的檢測技術。作為最前沿的刑事生物技術,DNA分析為法醫物證檢驗提供了科學、可靠和快捷的手段,使物證鑑定從個體排除過渡到了可以作同一認定的水平,DNA檢驗能直接認定犯罪、為兇殺案、強姦殺人案、碎屍案、強姦致孕案等重大疑難案件的偵破提供準確可靠的依據。隨著DNA技術的發展和套用,DNA標誌系統的檢測將成為破案的重要手段和途徑。此方法作為親子鑑定已經是非常成熟的,也是國際上公認的最好的一種方法。特別提到一點:同卵雙胞胎的DNA檢測結果是一樣的。
美國一位遺傳學研究者通過在網上發布的人類DNA信息,可以輕而易舉地確定從研究對象組中隨機選出的5個匿名者的身份,還找到了其整個家族,確定了近50人的身份。
在網上發布的遺傳數據,那些來自1000多人的長達幾十億個DNA字母的串子,看似是完全匿名的。但僅僅靠一些網上的聰明偵探手段,一位遺傳學研究者就把從研究對象組中隨機選出的5個人的身份確定了出來。不僅如此,他還找到他們的整個家族,確定了近50個人的身份,雖然這些親屬與研究一點也不沾邊。
這位研究者並未公布他所發現的人的姓名,但這項發表在周四的《科學》(Science)雜誌上的工作表明,保護參加醫學研究的志願者的隱私不是一個簡單的事情,因為他們提供的遺傳信息需要公開,以便科學家使用。
研究人員表示,“讓認為能夠完全保護隱私或使數據匿名的幻想繼續下去,已不再是一個可維持的立場。”
套用案例
1888年秋天,英國首都倫敦東區接連發生5起妓女遭殺害案件,多數受害人被開膛,但真兇一直未能確定。傳聞中的疑兇超過100人,甚至包括英國王室成員和首相。
2007年,迷戀研究此案的愛德華茲在一次拍賣會上買下一條帶有血跡的披肩,據稱為妓女凱薩琳·埃多斯兇殺案現場物品。
2014年9月7日,英國商人拉塞爾·愛德華茲和法醫學專家,藉助先進的法醫分析技術,成功破解困擾世人126年的謎:誰是英國連環殺手“開膛手傑克”。藉助分析和比對DNA樣本,認定波蘭美髮師阿倫·科斯明斯基為真兇。
科斯明斯基是猶太人,他被警方列為3名重點嫌疑人之一,一名目擊者也指認他為兇手。但是,警方沒有足夠證據指控科斯明斯基。他最終於53歲時死在精神病院。
愛德華茲鎖定了科斯明斯基。基因證據專家採用“真空吸取”的方式獲取了DNA樣本,與埃多斯後裔的DNA比對後,確定披肩上的血跡屬於埃多斯。
專家們還在披肩上的精液痕跡中發現了上皮細胞,並找到科斯明斯基妹妹的一名女性後代。比對顯示,DNA完全吻合。
基因工程
多活性多肽和蛋白質都具有治療和預防疾病的作用,它們都是從相應的基因中產生的。但是由於在組織細胞內產量極微,所以採用常規方法很難獲得足夠量供臨床套用。基因工程則突破了這一局限性,能夠大量生產這類多肽和蛋白質,迄今已成功地生產出治療糖尿病和精神分裂症的胰島素,對血癌和某些實體腫瘤有療效的抗病毒劑――干擾素,治療侏儒症的人體生長激素,治療肢端肥大症和急性胰腺炎的生長激素釋放抑制因子等100多種產品。
基因工程還可將有關抗原的DNA導入活的微生物,這種微生物在受免疫應激後的宿主體內生長可產生弱毒活疫苗,具有抗原刺激劑量大、且持續時間長等優點。
20世紀70年代創立的單克隆抗體技術在防病抗病方面雖然發揮了重要作用,但由於人源性單抗很難獲得,使得單抗在臨床上的套用受到限制。
抗生素在治療疾病上起到了重要作用,隨著抗生素數量的增加,用傳統方法發現新抗生素的幾率越來越低。為了獲取更多的新型抗生素,採用DNA重組技術已成為重要手段之一。
基因工程多肽、蛋白質、疫苗、抗生素等防治藥物不僅在有效控制疾病,而且在避免毒副作用方面也往往優於以傳統方法生產的同類藥品,因此引起了廣泛關注。
人類疾病都直接或間接與基因相關,在基因水平上對疾病進行診斷和治療,則既可達到病因診斷的準確性和原始性,又可使診斷和治療工作達到特異性強、靈敏度高、簡便快速的目的。於基因水平進行診斷和治療在專業上稱為基因診斷和基因治療。以補償失去功能的基因的作用,或是增加某種功能以利對異常細胞進行矯正或消滅。
在理論上,基因治療是治本治癒而無任何毒副作用的療法。不過,儘管至今國際上已有100多個基因治療方案正處於臨床試驗階段,但基因治療在理論和技術上的一些難題仍使這種治療方法離大規模套用還有一段很長的距離。不論是確定基因病因還是實施基因診斷、基因治療、研究疾病發生機理,關鍵的先決條件是要了解特定疾病的相關基因。隨著“人類基因組計畫”的臨近完成,科學家們對人體全部基因將會獲得全面的了解,這就為運用基因重組技術造逼於人類健康事業創造了條件。
不過,雖然基因技術向人類展示了它奇妙的“魔術師”般的魅力,但也有大量的科學家對這種技術的發展予以人類倫理和生態演化的自然法則的衝擊表示出極大的擔憂。從理論上來講,這種技術發展的一個極致就是使人類擁有了創造任何生命形態或從未有過的生物的能力。
發展計畫
人類基因組計畫(human genome project,HGP)是由美國科學家於1985年率先提出,於1990年正式啟動的。美國、英國、法蘭西共和國、德意志聯邦共和國、日本國和中國科學家共同參與了這一價值達30億美元的人類基因組計畫。這一計畫旨在為30多億個鹼基對構成的人類基因組精確測序,發現所有人類基因並確定其在染色體上的位置,破譯人類全部遺傳信息。與曼哈頓核子彈計畫和阿波羅登月計畫並稱為三大科學計畫。
2000年6月26日,參加人類基因組工程項目的美國、英國、法國、德國、日本和中國,六國科學家共同宣布,人類基因組草圖的繪製工作已經完成。最終完成圖要求測序所用的克隆能忠實地代表常染色體的基因組結構,序列錯誤率低於萬分之一。95%常染色質區域被測序,每個Gap小於150kb。完成圖將於2003年完成,比預計提前2年。
美國和英國科學家2006年5月18日在英國《自然》雜誌網路版上發表了人類最後一個染色體——1號染色體的基因測序。
在人體全部22對常染色體中,1號染色體包含基因數量最多,達3141個,是平均水平的兩倍,共有超過2.23億個鹼基對,破譯難度也最大。一個由150名英國和美國科學家組成的團隊歷時10年,才完成了1號染色體的測序工作。
科學家不止一次宣布人類基因組計畫完工,但推出的均不是全本,這一次殺青的“生命之書”更為精確,覆蓋了人類基因組的99.99%。解讀人體基因密碼的“生命之書”宣告完成,歷時16年的人類基因組計畫書寫完了最後一個章節。
作為人類基因組計畫的後續計畫,The ENCODE Project 在2003年9月啟動的跨國研究項目。該項目旨在解析人類基因組中的所有功能性元件。該項目聯合了來自美國,英國,西班牙,新加坡和日本的32個實驗室的422名科學家的努力,獲得了迄今最詳細的人類基因組分析數據(他們獲得並分析了超過15兆兆位元組的原始數據)。研究花費了約300年的計算機時間,對147個組織類型進行了分析,以確定哪些能打開和關閉特定的基因,以及不同類型細胞之間的“開關”存在什麼差異。
2012年9月5日,ENCODE項目的階段性研究結果被整理成30篇論文發表於《自然》(6篇),《基因組研究》(6篇)和《基因組生物學》(18篇)上。研究結果顯示,人類基因組內的非編碼DNA至少80%是有生物活性的,而並非之前認為的“垃圾” DNA (junk DNA)。這些新的發現有望幫助研究人員理解基因受到控制的途徑,以及澄清某些疾病的遺傳學風險因子。
2012年12月21日,ENCODE項目被《科學》雜誌評為本年度十大科學突破之一。
2012年12月28日,早老素同源蛋白PSH的晶體結構。
轉基因食品相關知識
| 轉基因食品(Genetically Modified Foods,GMF)是利用現代分子生物技術,將某些生物的基因轉移到其他物種中去,改造生物的遺傳物質,使其在形狀、營養品質、消費品質等方面向人們所需要的目標轉變。以轉基因生物為直接食品或為原料加工生產的食品就是“轉基因食品”。 |
