
概述:
 肌球蛋白分子
肌球蛋白分子肌球蛋白是一種馬達蛋白(motor protein),由Kuehne於1864年在研究骨骼肌收縮時發現並命名[1]。肌球蛋白是一種超家族的蛋白質,共分為11類,其中10類為非傳統肌球蛋白(unconventional myosin),另一類肌球蛋白Ⅱ稱為傳統肌球蛋白(conventional myosin)[2]。本文對肌球蛋白Ⅱ(全文簡稱肌球蛋白)進行綜述,對近年來在肌球蛋白結構、性質、功能研究上取得的新成就給予介紹。
肌球蛋白的結構
肌球蛋白是長形不對稱分子,形狀如“Y”字,長約160nm。電子顯微鏡下觀察到它含有兩條完全相同的長肽鏈和兩對短肽鏈,組成兩個球狀頭部和一個長桿狀尾部。肌球蛋白分子量約460kD,長肽鏈的分子量約240kD,稱重鏈;短鏈稱輕鏈。將肌肉肌球蛋白用5,5′-二硫雙(α-硝基苯甲酸,DTNB)處理後放出的一對輕鏈,稱為DTNB鏈,分子量約18kD;另兩條輕鏈只有在鹼性(pH 11.4)條件下才能分離出來,稱鹼性輕鏈,分子量分別為25kD和16kD。非肌細胞如粘菌的肌球蛋白的形狀和結構與肌細胞的肌球蛋白非常相似,但它不存在DTNB鏈,兩對不同的輕鏈稱必需輕鏈(essential light chain)和調節輕鏈(regulatory light chain),分子量分別為16kD和18kD[2]。閻隆飛等於1963年首次從菸草微管束中分離出肌動球蛋白,證明其具有ATP酶活性,這是高等植物中存在肌動蛋白和肌球蛋白的第一個證據[3]。馬永澤和閻隆飛(1989)證明,豌豆葉片的卷鬚中存在肌球蛋白,SDS-PAGE測得兩條重鏈的分子量為165kD,兩對輕鏈的分子量分別為17kD和15kD[4]。
 肌球蛋白分子
肌球蛋白分子在肌球蛋白超家族中,不管其來源如何,其頭部區域都有相當高的同源性,特別是ATP和肌動蛋白的結合位點非常保守。兩條重鏈的氨基末端分別與兩對輕鏈結合,形成兩個球狀的頭部和頸部調節結構域,稱為S1(subfragment 1),餘下重鏈部分組成肌球蛋白長桿狀的尾部(圖1)。在一定條件下,胰凝乳蛋白酶能把肌球蛋白切為兩部分,帶有兩個頭部的部分稱為重酶解肌球蛋白(heavy meromyosin,HMM),另一部分叫輕酶解肌球蛋白(light meromyosin,LMM)。重酶解肌球蛋白的尾部稱為S2(subfragment 2)。肌球蛋白N末端的頭部S1為馬達功能區,在離體條件下,單獨的S1就能依賴其ATP酶活性產生力,從而驅使肌動蛋白絲運動,只是滑動速度比全長的肌球蛋白慢,可見肌球蛋白表現最佳馬達活性需要S2。肌球蛋白的尾部是超螺鏇結構,其胺基酸序列由典型的捲曲的捲曲型α螺鏇(coiled-coiled α-helix)組成。所有肌球蛋白在輕鏈結合下方都有一個脯氨酸殘基,根據其保守性和結構特性,此脯氨酸殘基被定義為頭部和尾部連線點(head/tail junction)。肌球蛋白以聚合形式參與細胞生理過程,單個的肌球蛋白分子沒有功能。肌球蛋白通過組裝域自我組裝成具有雙極性的粗絲(thick filament),對其實現合適的功能至關重要。1993年,Rayment等通過甲基化修飾肌球蛋白S1的賴氨酸殘基獲得了高質量S1的晶體,在2.8A高解析度下解析了雞胸肌肌球蛋白三維空間結構,後來又結晶了盤基網柄菌肌球蛋白S1,並證明S1的ATP結合袋(ATP binding pocket)水解ATP時,有較大的構象變化,它為設計突變體提供了極有價值的理論依據,是肌球蛋白研究史上的新突破。
肌球蛋白分子圖解
肌球蛋白的性質
肌球蛋白屬球蛋白類,不溶於水而溶於0.6mol/ml的KCl或NaCl溶液。它具有酶活性,通過與肌動蛋白相互作用,水解ATP的末端磷酸基團,同時也能水解GTP、CTP等,將化學能轉化為機械能,從而產生各種形式的運動。物理化學研究表明,肌球蛋白溶液加入ATP後,其粘度和流動雙折射顯著下降。後來證實這是由於肌動蛋白與肌球蛋白複合物的分解,形成兩種軸比較小的蛋白質分子而引起。因此,加入ATP前後的粘度變化是鑑定肌球蛋白製備物中是否存在肌動蛋白最簡單可行的方法。骨骼肌肌球蛋白ATP酶在Mg2+存在時活性很低,但在K+及EDTA或Ca2+存在時活性可增加10倍以上。
 肌球蛋白分子
肌球蛋白分子植物細胞中的肌球蛋白含量很少,只有肌肉含量的1%,從植物中純化肌球蛋白非常困難,故植物肌球蛋白研究的進展落後於動物和微生物。馬永澤和閻隆飛(1989)研究表明豌豆葉片卷鬚中的肌球蛋白的ATP酶活性可被兔肌F-肌動蛋白激活5.6倍[4]。劉熊和閻隆飛從玉米花粉中純化到的肌動蛋白可激活兔肌肌球蛋白ATP酶活性7倍,他們用微量測熱法證明兔肌肌球蛋白的頭部S1和HMM片段在ATP存在和缺乏時表現出的熱力學性質相同。
肌球蛋白的功能
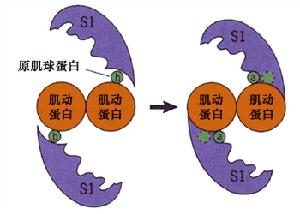
肌球蛋白作為細胞骨架的分子馬達,是一種多功能蛋白質,其主要功能是為肌肉收縮提供力。纖絲滑動學說(sliding filament theory)認為肌肉收縮是由於肌動蛋白細絲與肌球蛋白絲相互滑動的結果。在肌肉收縮過程中,粗絲和細絲本身的長度都不發生改變,當纖絲滑動時,肌球蛋白的頭部與肌動蛋白的分子發生接觸(attachment)、轉動(tilting),最後脫離(detachment)的連續過程,其結果使細絲進行相對的滑動。此學說當時有強有力的實驗依據,但沒有在分子水平加以證實。
關於肌絲滑動的分子機理,一直是近半個世紀來十分活躍的研究領域。具有很大影響的經典擺動橫橋模型(classic swinging crossbridge model)至今仍被教科書所採用。但隨著研究手段不斷創新,取得的許多結果已不能用擺動橫橋模型進行解釋,便產生了新的肌絲滑動學說,具有代表性和影響力的是擺動桿臂模型(swinging ever arm model)和線形馬達模型(linear motor model)學說
圖2 肌球蛋白產生力的三個代表模型[6]
經典的擺動橫橋模型強調,肌絲滑動時肌球蛋白頭部與頸部相對位置沒有改變,頭部首先以90度角與肌動蛋白絲結合,當ATP水解時,肌球蛋白頭部傾斜45度,使肌動蛋白絲產生一定的移動,然後頭部與肌動蛋白絲脫離,分子回到原來狀態。擺動桿臂模型則認為,肌絲滑動時,肌球蛋白頭部與頸部的相對位置發生一定的改變,但頭部與肌動蛋白絲的結合角不變,頸部相當一根槓桿(lever),通過分子屈伸使肌動蛋白絲移動。但用它來解釋肌球蛋白驅使肌動蛋白絲滑動的步幅(step size)時遇到了困難。線性馬達學說提出,肌球蛋白頭部象一個馬達,肌動蛋白絲沿其直線滑動。它可解釋大運動步幅的產生,但沒有直接的實驗依據。不同來源的肌球蛋白驅動肌動蛋白運動速度有很大的差異,骨骼肌肌球蛋白、平滑肌肌球蛋白和盤基網柄菌肌球蛋白推動肌動蛋白運動速度分別為8.4μm/s、1μm/s和3μm/s。還有人從藻類細胞(Chara)純化到一種肌球蛋白,能以50μm/s的速度驅動肌動蛋白運動。至今尚無一種比較完善的、能普遍被人們接受的理論以解釋肌絲滑動的原理。
肌球蛋白也廣泛存在於非肌細胞中,它是細胞骨架的組成成分,為細胞質流動、細胞器運動、物質運輸、有絲分裂、胞質分裂和細胞的頂端生長等提供所需的力,參與細胞的吞噬、運動、受精和吸收等生理過程,充當非肌細胞生命活動的不同層次的調節者,從簡單的細胞間的信號傳遞到指導向化性遷移和細胞形狀的改變等較高級的調節。研究表明,肌球蛋白為盤基網柄菌在振盪培養下正常生長和分裂、表面蛋白受體成帽(surface receptor capping)、多細胞發育成子實體(fruiting body)、細胞與器壁的脫粘附(adherence)等生命活動所必需[6]。閻隆飛等在80年代的研究結果表明,肌球蛋白在高等植物如黃瓜等花粉管伸長和豌豆葉片卷鬚運動中可能執行重要功能。Kinkema等(1994)證明植物肌球蛋白在植物生長發育的不同時期及不同部位有不同的生理功能[9]。
近半個多世紀來,肌球蛋白的研究日益熱門,但其產生力的分子機理一直未能闡明。最大的困難和限制因素之一是,直至80年代初,人們還不能對肌球蛋白與肌動蛋白相互作用所產生力的大小進行體外測試。1986年,Spudich實驗室才首次建立了體外模擬滑動體系(in vitro motility assay)[10]。而Finer等用雙光鉗成功地進行單分子運動測試已是1994年的報導了[11]。人們能在單分子水平上研究肌絲的相互滑動,測量產生力的大小,終於使肌球蛋白研究進展上了一個新台階。
肌球蛋白與心臟功能
心臟收縮-舒張是一個非常複雜的生理過程,受諸多生理性和/或病理性因素影響而發生變化,因此而影響心功能。尤其臨床上許多疾病都伴有心功能改變,嚴重時出現心功能障,心肌收縮力下降,心輸出量減少。
隨著分子生物學等相關學科的迅猛發展,人們從細胞水平、分子水平對心肌收縮-舒張過程及其調節的諸多參與成分各自的作用及相互間作用有了更進一步的了解和認識。近十幾年來,人們針對糖尿病、甲狀腺功能異常(包括功能亢進和低下)、心肌肥厚、心肌病、缺氧等病理條件下引起的心功能改變,特別是收縮蛋白、調節蛋白與心功能的關係做了大量深入細緻的工作。
1 收縮蛋白和調節蛋白
收縮蛋白包括肌球蛋白和肌動蛋白。肌球蛋白是由學者Kuhne於1859年首先報導的,半個多世紀之後,對肌球蛋白的生化分析才開始進行。肌球蛋白是心肌粗肌絲的主要成分,分子呈桿狀,一端具有兩個球形區域,似豆芽的頭部,由兩條重鏈(MHC)和兩對輕鏈(MLC)構成,是肌球蛋白重要生物活性所在地,另一端是一個絲狀“尾巴”,由兩股α-螺鏇肽鏈絞在一起形成一種盤卷螺鏇結構[1]。肌球蛋白具有二個生物學作用:一是具有ATP酶活性,能裂解ATP,釋放化學能;二是具有與肌動蛋白結合的能力。研究表明心臟的MHC是由兩種基因編碼,即α-MHC和β-MHC基因,這些基因產物在肌球蛋白分子中形成二聚體,所以相應的有三種分子異構體存在,即V1(α、α同源體)、V2(α、β異源體)、V3(β、β同源體)。由於α、β-MHCATP酶活性不同,因此不同的異構體之間所具有的ATP酶活性及收縮活性也不同。肌球蛋白ATP酶活性主要由心肌所含V1或V3的量多少而決定,故肌球蛋白以V1占優勢的心肌ATP酶活性最高,肌肉收縮速率最快,耗能也最多,而以V3占優勢的心肌情況正相反,以V2占優勢的心肌表現介於兩者之間[2,3]。肌球蛋白異構體之間的轉換是心肌的適應性改變,是心臟本身負荷和能量供應兩方面調節適應的結果。V1通過增加心肌收縮速度來增加供能達到能量供求平衡,V3通過減少耗能而適應壓力超負荷。當能量供不應求時,肌球蛋白異構體向V3轉化,使ATP酶活性下降,心肌收縮功能降低,表現為Vmax下降,最大張力正常,而達到最大張力的時間延長,心肌作功時耗氧量下降,結果使心臟在節能的情況下產生同樣的張力,所以V3增加雖可使心肌速度變慢但是卻提高了機械效率。正常哺乳動物和人的心室肌球蛋白異構體的分布與種屬、年齡等因素有關。成年人左心室心肌肌球蛋白以V3為主占60%~90%,而小哺乳類動物左心室心肌肌球蛋白以V1為主占60%~90%,人類和哺乳類小動物心房肌球蛋白以V1為主[4]。
對心肌肥厚等病理狀態研究顯示,心臟肌球蛋白基因表達及蛋白異構中存在著可塑性,推測這可能是動物機體的一種適應反應,例如超負荷刺激引起大鼠心肌肥厚可誘導左心室β-MHC基因表達及V3肌球蛋白增多,結果使心肌耗氧降低,收縮速率下降,被認為是一種經濟的適應性反應[5]。
與肌球蛋白相比,肌動蛋白結構及功能相對簡單。分子單體為球形,單體上有與肌球蛋白頭相結合的位點,許多單體相互連線形成兩條有極性的相互纏繞螺鏇體。
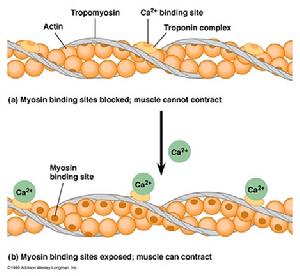
調節蛋白包括原肌球蛋白(Tm)和肌鈣蛋白(Tn),Tm和Tn結合鈣離子構成調節蛋白複合物,通過影響肌動蛋白和肌球蛋白之間的相互作用調節收縮活動[6]。Tm分子由二條完全相同或不同的螺鏇形肽鏈組成(同源或異源二聚體),不同組織來源(如房、室)和特殊種類的Tm不盡相同。Tn由三種亞單位組成,即TnI、TnT、TnC。TnI是肌動球蛋白複合物的Tn抑制形,具有調節肌動蛋白和肌球蛋白相互作用的能力,這種調節作用主要是一種抑制作用。TnT即肌鈣蛋白結合原肌球蛋白,其作用是將肌鈣蛋白複合體附著在Tm上。TnC即鈣結合肌鈣蛋白,是鈣離子的受體,具有兩個高親和力和兩個低親和力的鈣離子結合點。這三種肌鈣蛋白亞型以協同方式相互之間與Tm及肌動蛋白互相作用[7]。
2 病理條件肌球蛋白的改變
2.1 機械性作用
利用乳鼠分離培養的心肌細胞,在體外進行機械牽張與心肌肥厚的實驗研究表明,機械牽張引起大鼠心肌細胞MHCmRNA表達增高。Shyu等人[8]用Northernblot分析研究了分離培 養的乳鼠心肌細胞周期性牽張對MHCmRNA表達的影響,與對照組相比顯示在不同時相點MHCmRNA最高增加了12倍。Vandenburgh等人[9]研究證實機械作用增加MHC含量,包括β-MHC和α-MHC都增加。Kojima等人[10]在自發性高血壓(SHRs)動物模型研究中看到從9周齡到25周齡經vehicle治療組的SHRs,MHCV3異構體進行性增加。
2.2 限制飲食、缺氧
限制飲食使大鼠心肌V3肌球蛋白異構體增高[11],早已被人們所認識[12],這些改變被認為由血中T3水平降低而觸發[13],而最近一些研究顯示限制飲食使β-MHC增高並無循環T3的明顯改變[12],Pissarek等也得出相同的結果。但Swoap等認為這不能排除由於心臟甲狀腺激素受體的減少而引起一個功能性高甲狀腺素的可能性[14]。Pissarek等在慢性低氧(CHH)大鼠模型研究中發現大鼠左、右心室都有一個明顯的α-MHC向β-MHC的轉換。慢性缺氧引起心肌收縮器(Apparatus)實質性變化,V3肌球蛋白異構體的表達使得心臟降低收縮耗能,提高工作效率[15]。
3 幾種疾病狀態下肌球蛋白及調節蛋白改變
3.1 糖尿病
糖尿病是許多常見的慢性病之一,它與心血管疾病引起的死亡有較密切的關係。其心功能的破壞不依賴於血管性疾病,提示在糖尿病存在一個原發的心肌的缺陷。大量動物實驗研究提示慢性糖尿病動物心功能改變與收縮蛋白ATP酶活性受抑制及肌漿網(SR)和肌膜(SL)鈣運輸異常有關,這些異常經胰島素治療都能逆轉。另外用鈣通道阻滯劑維拉帕米治療,心臟收縮蛋白ATP酶活性及肌漿網鈣離子活性得到改善[16~18]。進一步研究顯示慢性糖尿病動物心臟肌球蛋白異構體-βMHC與ATP酶活性降低和收縮速率(Shortingvelocity)密切相關[19]。
在STZ(Streptozotocin)糖尿病大鼠心肌力學和肌球蛋白酶學研究中,Takeda等人報導了糖尿病影響心肌收縮,肌球蛋白V1向V3轉換及心臟能量學的變化。與心肌肌球蛋白以β形占優勢相比較,肌球蛋白以α重鏈占優勢的心肌表現收縮速率增加,高ATP酶活性,收縮能量消耗也增加。
在齧齒動物心臟一些病理條件下如高血壓心肌肥厚、糖尿病、心肌梗死及老齡心肌,肌球蛋白異構體也顯示出明顯的轉換。糖尿病心臟收縮速率的降低也許可用大鼠模型中肌球蛋白異構體的變化得到部分或全部解釋[20]。
成年人心室肌球蛋白以V3異構體占優勢,所以這也許是嚴重病理狀態下人類心臟中並沒有觀察到肌球蛋白ATP酶活性變化的原因,但觀察到肌纖維ATP酶曲線下降,推測病理狀態下微小差異可能存在於人類肌球蛋白重鏈,這種差異用一個範圍的焦磷酸鹽凝膠電泳往往觀察不到。故認為非常小的異構酶(Isoenzyme)轉換與收縮蛋白的其它主要改變一起可能導致肌纖維活性的明顯變化[21]。
除了收縮蛋白,調節蛋白及鈣離子因素也直接或間接影響心臟的功能。在脊椎動物的橫紋肌,細肌絲的調節成分TnTm負責傳導收縮蛋白活動中鈣離子效應,並且當鈣離子缺乏時則抑制這種活動[22]。人們通過對照組或糖尿病組調節複合物TnTm存在的條件下利用對照組或糖尿病組肌球蛋白觀察Ca2+依賴性心臟肌動球蛋白ATP酶活性。當糖尿病大鼠心臟的肌球蛋白被從對照組心臟分離的調節蛋白複合物調節時,肌球蛋白ATP酶可以部分逆轉,提示糖尿病大鼠病理模型中調節蛋白能部分上調心臟的肌球蛋白[23]。在SDS平板凝膠中,來自慢性糖尿病大鼠和對照組動物心臟的調節蛋白在TnI和TnT表現不同,加了對照組動物TnTm的糖尿病心臟被調節的肌動球蛋白ATP酶活性發生逆轉,這可能說明糖尿病心肌病理中或TnTm亞單位出現的量不同或存在調節蛋白亞單位異構酶組成不同。另外來自糖尿病心臟的調節複合物被重組,導致心臟肌動球蛋白系統調節中鈣敏感性下降[24]。而對來自正常的和疾病的人類心肌完整的和分離的心肌中PKC活性作用的一些研究中,Gwathmey和Hajjar報導了肌絲的Ca2+敏感性和收縮活性的改變可能是由TnI和TnT磷酸化所致[25]。也有報導肌球蛋白輕鏈-2(MLC-2)與PKC的直接磷酸化或PKC的受體介導的刺激物都分別可以導致去膜的[skinned]心肌細胞和肌纖維中Ca2+敏 感性和ATP酶活性的增加[26]。
3.2 甲狀腺功能亢進或低下
心臟是甲狀腺激素作用的一個主要靶器官,激素水平過高或過低時都明顯影響心臟功能。T3對心臟功能的影響是通過它對心臟的直接作用和間接作用。直接作用指T3對心臟細胞的直接效應,是通過核或核外機制介導的。核外T3效應:它的發生不依賴與核T3受體的結合,增加蛋白質合成,主要影響胺基酸、糖及鈣離子的穿膜運輸;核的T3效應:通過T3與特殊的核受體蛋白結合而被介導,導致T3反應性心臟基因轉錄增加,另外T3對肌膜(SL)Ca2+a-ATP酶也存在核外效應,引起鈣離子從肌細胞流出增加。甲狀腺激素對蛋白合成的效應使總體心臟蛋白形成增加,並且增加特殊蛋白的合成率,這種增加遠遠超過了由甲狀腺激素誘導的蛋白合成的一般性增加。對其它一些特殊的蛋白象肌球蛋白重鏈(MHC)β合成率是降低的,使肌球蛋白V1異構體增加,V3降低,心肌收縮速率增加,ATP消耗也增加。間接作用是T3引起外周改變進一步導致血液動力學變化而影響心功能[27~29],甲亢時,T3除了增加V1肌球蛋白異構體,也增加心肌的舒張率,被認為與肌漿網(SR)的Ca2+-ATP酶泵的活性有關。有學者報導T3誘導增加了SRCa2+-ATP酶(SERCA2)基因的轉錄使SERCA2mRNA水平增加[30]。SERCA2是一個離子泵,負責舒張期將胞漿中的鈣離子運回到肌漿網。V1肌球蛋白異構體和SERCA2泵數量增加導致甲亢時心臟ATP消耗明顯增加,另外ATP化學能少部分用於心肌收縮過程,大部分用於產生熱量,降低了心臟的收縮效率[31]。
甲狀腺功能低下時情況正相反。T3缺乏在成年嚙齒類動物心臟觀察到肌球蛋白異構體V1向V3轉換,SERCA2活性降低。由此而引起心臟收縮率,代謝等方面的改變在此不再贅述。
總之在病理條件下,肌球蛋白異構體明顯轉化,肌球蛋白由V1向V3轉化,引起心肌收縮力下降,收縮速度變慢。耗氧降低被認為是心肌的一種適應性反應。另外調節蛋白,SERCA2等結構的改變從不同作用環節影響心臟的功能。
參考資料:
http://www.wanfangdata.com.cn/qikan/periodical.Articles/smdhx/smdh99/smdh9902/990208.htm
http://www.daifumd.com/_daifumd/blog/html/678/article_85789.html
