
 糖皮質激素受體作用
糖皮質激素受體作用糖皮質激素受體(glucocorticoid reccptor,GR)屬於保守的核受體超家族中的一員,這個家族中包括鹽皮質激素,雄激素、雌激素、黃體酮、甲狀腺激素、維A酸、維生素D等多種激素的受體。
在激素結合的狀態下,這些受體能夠特異性的與靶基因的啟動子結合併調節這些啟動子的活性,因此,這些受體又被稱為配基依賴性的轉錄因子。在細胞水平上,大多數己知的糖皮質激素(glucocorticoids,GC)的作用都是由一種94KD的細胞內蛋白,糖皮質激素受體介導的。
結構
糖皮質激素受體的一級結構可分為N末端和C末端及3個獨立的功能區。N末端和C末端的轉錄激活功能區AF-1和AF-2可控制靶基因轉錄。例如Hittelman等研究發現GR AF-1能夠募集到使糖皮質激素受體轉錄增加或減少的調節因子,如維生素D受體相互作用蛋白DRIP150和DRIP205以激素依賴的方式與糖皮質激素受體配體結合域相互作用並促進糖皮質激素受體轉錄激活。
 糖皮質激素受體
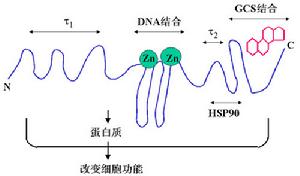
糖皮質激素受體糖皮質激素受體蛋白質結構3個獨立的功能區包括:激素結合區(hormone-binding domain,HBD),DNA結合區(DNA-binding domain,DBD)和免疫原區(immunogenic domain,ID)。其中HBD位於COOH端,主要功能包括:①與激素配體結合,參與糖皮質激素受體二聚體形成;②可與熱休克蛋白(hot shock protein,HSP)結合;③其序列內含有其他轉錄因子如AP-1的作用位點及核轉位信號的核定位序列。DBD位於糖皮質激素受體蛋白的中部,負責與靶基因啟動子或增強子上的糖皮質激素反應元件(glucocorticoids response element,CRE)結合;ID位於NH2端,具有特異性抗原活性,含有多個可磷酸化的絲氨酸和蘇氨酸位點,與糖皮質激素受體的磷酸化和脫磷酸化有關,並參與糖皮質激素受體的反式激活作用。
Wang等研究發現,在無激素存在時,糖皮質激素受體的Ser203位點易被磷酸化,在激素存在條件下,Ser203位點和Ser211位點均被磷酸化,糖皮質激素受體的Ser203磷酸化形式主要在胞質,而Ser211磷酸化形式主要在核內。這提示不同的磷酸化受體種類定位於獨特的亞細胞器,可能調節受體功能的不同方面。
糖皮質激素受體結構穩定是保證其發揮正常生理調節功能的必要保證,故糖皮質激素受體本身結構的改變可直接影響到其轉錄功能。Nagano等在對白血病細胞中GC抵抗的研究中發現如果糖皮質激素受體的半胱氨酸643被精氨酸取代後,其與類固醇的親和力和轉錄活性均降低,這就大大影響了糖皮質激素受體的調節功能。
分類
糖皮質激素受體包括糖皮質激素受體α和糖皮質激素受體β,兩者為糖皮質激素受體基因同一轉錄產物通過不同的剪下方式剪下的結果。
 糖皮質激素受體
糖皮質激素受體研究表明:糖皮質激素受體α和糖皮質激素受體β的前727個胺基酸完全相同,從第728個胺基酸開始糖皮質激素受體α有50個胺基酸序列,而糖皮質激素受體β只有15個胺基酸序列。在mRNA 水平上,兩者都包含第1到第8外顯子,不同之處為糖皮質激素受體α包含9α外顯子,而糖皮質激素受體β包含9β外顯子。糖皮質激素受體α在結構上包括3個功能區,即氨基端的轉錄活化區、羧基端的糖皮質激素結合區和中間的DNA結合區。因為糖皮質激素受體α在幾乎所有組織和細胞中均有表達,在絕大多數細胞中其含量也遠遠超過糖皮質激素受體β,且糖皮質激素(GC)主要通過結合糖皮質激素受體α發揮作用。因此對糖皮質激素受體α的表達、生化特點及生理功能等早就有了詳細的研究,對糖皮質激素受體β的研究最近幾年才引起關注,研究發現糖皮質激素受體β對糖皮質激素受體α的功能有拮抗作用,可能對糖皮質激素的生理及藥理功能起著重要的負性調節作用。
GR與GC
糖皮質激素廣泛用於各種疾病的治療,特別是對某些免疫性及炎症性疾病有很好的療效,是治療首選藥物,但長期大量套用糖皮質激素可以導致高血壓、糖尿病、骨質疏鬆等不良反應。不同患者對糖皮質激素治療的敏感性不同,特別是有部分患者對糖皮質激素治療不敏感,盲目增加劑量不僅不能帶來預期的療效,反而引起嚴重不良反應,甚至導致患者死亡。因此當需要長期大劑量使用糖皮質激素時,我們必須考慮如何以最小劑量取得理想療效並最大限度減少不良反應。
 糖皮質激素
糖皮質激素糖皮質激素受體是糖皮質激素髮揮療效的關鍵環節,糖皮質激素受體α的密度及其與糖皮質激素的親和力與糖皮質激素的療效呈正相關,糖皮質激素受體β則對糖皮質激素功能起重要負性調節因素。如何根據患者糖皮質激素受體的表達情況調節糖皮質激素的使用劑量,如何干擾受體後過程,在阻斷不良反應的同時保持療效,如何恢復糖皮質激素耐藥患者對糖皮質激素治療的敏感性等正成為研究的熱點,這可能將在糖皮質激素用藥個體化、增強療效和減少不良反應方面帶來重大突破。
許多臨床研究表明,糖皮質激素受體α數量及其與糖皮質激素的親和力與糖皮質激素的療效密切相關。如Cho等通過放射免疫法測定外周血單個核細胞( PBMC)糖皮質激素受體α與糖皮質激素的親和力,發現在老年嚴重哮喘患者中, 糖皮質激素治療不敏感患者糖皮質激素受體α與糖皮質激素的親和力顯著低於糖皮質激素治療敏感患者及正常對照。Pujols等研究了間質性肺病患者肺組織糖皮質激素受體α密度與糖皮質激素療效的關係,發現糖皮質激素治療不敏感患者糖皮質激素受體α密度顯著低於糖皮質激素治療敏感患者。
關於糖皮質激素受體β與糖皮質激素療效的關係,最近幾年有一些研究,結果不一,多數研究表明,糖皮質激素受體β對糖皮質激素的療效起負性調節作用。Oakley等用h糖皮質激素受體α、h糖皮質激素受體β、小鼠乳房腫瘤病毒細胞- 螢光素酶報告基因構建PHHluc質粒,然後轉染SV40轉化的非洲綠猴腎細胞(COS - 1 細胞),所有細胞中糖皮質激素受體α的轉染量相同,而糖皮質激素受體β的轉染量則分別為糖皮質激素受體α轉染量的0倍、5倍、10倍,將該細胞與糖皮質激素共同培養,通過測定螢光素酶的活性了解糖皮質激素受體α的功能。結果培養18 h後,含糖皮質激素受體β5倍、10倍的非洲綠猴腎細胞- 1和COS -1細胞中, 糖皮質激素受體α的活性分別下降3611%和6317% ,這表明糖皮質激素受體β可以抑制糖皮質激素受體α的功能。
臨床研究方面,Sousa等研究發現糖皮質激素治療不敏感支氣管哮喘患者炎症細胞上糖皮質激素受體β的密度顯著高於激素治療敏感患者;Honda等則用RT2PCR方法測定潰瘍性結腸炎患者及正常對照人群中PBMC 上糖皮質激素受體βmRNA的表達情況,在一定的擴增條件下,12例對糖皮質激素治療不敏感的患者中10例糖皮質激素受體βmRNA陽性,11例對糖皮質激素治療敏感的患者中則僅有1例陽性,正常對照組20 例中2 例陽性。Chikanza的研究也表明糖皮質激素治療不敏感類風濕性關節炎患者PBMC上糖皮質激素受體β的表達明顯增多。但也有一些研究表明,糖皮質激素受體β的表達量與糖皮質激素治療不敏感之間無相關性。如Gagliardo等用巢式PCR方法測定PBMC上糖皮質激素受體βmRNA的表達情況,其中14例為對糖皮質激素治療依賴的支氣管哮喘患者、8例為未用糖皮質激素治療的活動期支氣管哮喘患者、11例為穩定期未用藥的支氣管哮喘患者及8例正常對照人群。結果發現糖皮質激素受體β的表達量在4組人群中差異無統計學意義。
GC生理藥理作用
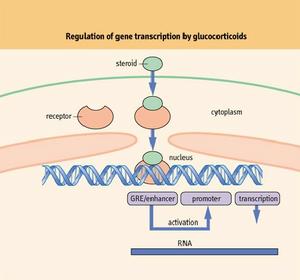
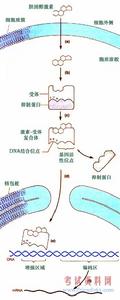
糖皮質激素主要通過經典基因組效應產生藥理作用,在無糖皮質激素存在的情況下,糖皮質激素受體α以受體複合物的形式存在於細胞漿中,該受體複合物包括受體多肽、兩個熱休克蛋白(HSP)90及其他一些蛋白成分,這些蛋白成分阻止受體多肽在游離狀態下與核內DNA產生相互作用。當糖皮質激素受體與糖皮質激素結合後,HSP 90的構象發生改變,糖皮質激素受體α2糖皮質激素複合物與HSP 90等蛋白成分分離並進入細胞核,在細胞核內與激素應答基因啟動區的激素反應元件(GRES)結合,從而導致相應基因的轉錄活性增強併合成相應的蛋白質,脂皮質素-1、黏附分子、核酸內切酶、神經內肽酶、血管緊張素轉化酶等,產生生理及藥理作用。如脂皮質素-1可以抑制磷脂酶A2的代謝,從而影響花生四烯酸級聯,抑制炎症介質的合成,在糖皮質激素抗炎作用中發揮重要作用。糖皮質激素不僅可以使某些基因的轉錄活性增強,
 糖皮質激素與DNA
糖皮質激素與DNA研究表明糖皮質激素受體α主要通過和活化蛋白-1(AP - 1)、核因子-κB(NF -κB)等轉錄因子相互作用,從而抑制以上這些基因的轉錄。此外,糖皮質激素受體α還可以通過受體後效應產生作用,如影響mRNA的穩定性、影響翻譯及分泌過程等。體外實驗表明,當糖皮質激素的用量達到相當於潑尼松250 mg時,體內糖皮質激素受體α即達到飽和狀態,理論上繼續增加劑量對療效應該無影響,但臨床實踐表明,超大劑量糖皮質激素特別是衝擊療法對某些嚴重疾病有顯著療效。
目前認為超大劑量糖皮質激素除了有經典的基因組效應外,還可能通過非基因組效應產生作用,非基因組效應包括經典非基因組效應和非經典非基因組效應,經典非基因組效應可能主要通過細胞膜上的糖皮質激素受體介導,而非經典非基因組效應可能主要通過與細胞膜的直接物理化學反應介導,非基因組效應起效時間短、產生療效快,具體機制尚未完全清楚。一般認為當糖皮質激素的用量小於相當於潑尼松250 mg時,非基因組效應很小,主要通過基因組效應產生臨床療效。
GR與細胞因子
對糖皮質激素治療不敏感的患者糖皮質激素受體α、糖皮質激素受體β的表達異常大部分是獲得性的,研究表明,細胞因子對糖皮質激素受體α、糖皮質激素受體β的表達及功能起重要的調節作用。
 細胞凋亡
細胞凋亡Kam等的研究發現IL - 2和IL - 4聯合作用可以使人PBMC上糖皮質激素受體α的密度增加,但其與糖皮質激素的親和力則顯著下降。Sher等的研究得出類似的結果。他們測定了17例對糖皮質激素治療不敏感的哮喘患者PBMC上糖皮質激素受體α的密度與親和力,發現15例親和力低下,將PBMC置體外培養48 h,其親和力可以恢復正常,但如果在培養基中同時加入IL - 2 和IL - 4,則仍保持低親和力。研究表明,IL - 2和IL - 4的聯合作用可以顯著降低人PBMC上糖皮質激素受體α與糖皮質激素的親和力,對糖皮質激素的生理及藥理功能起負性調節作用。Spahn等則研究了其他細胞因子對糖皮質激素受體α親和力的影響,他們發現IL - 13 也可以降低糖皮質激素與糖皮質激素受體α的親和力,IL - 1、IL - 3、IL - 5、IL - 7、IL - 8、IL - 12和GM-CSF則對糖皮質激素受體α與糖皮質激素的親和力無顯著影響。Spahn等在體外培養的Helas3 細胞中加或不加TNF - α,24 h 後用RT2PCR 方法測定糖皮質激素受體α、糖皮質激素受體β mRNA 的表達量,結果發現TNF - α可以使糖皮質激素受體αmRNA的數量增加115倍,使糖皮質激素受體βmRNA的數量增加210 倍。用Western 印跡方法測定糖皮質激素受體α、糖皮質激素受體β蛋白的表達情況時得出類似的結果,TNF -α可以使糖皮質激素受體α的數量增加115倍,使糖皮質激素受體β的數量增加315 倍。該試驗表明TNF -α可以增加糖皮質激素受體α、糖皮質激素受體β蛋白的表達數量,且糖皮質激素受體β的增幅顯著大於糖皮質激素受體α。同樣的試驗表明IL - 1也有類似的作用。有學者用免疫組化法測定糖皮質激素治療不敏感支氣管哮喘患者、糖皮質激素治療敏感的支氣管哮喘患者及正常人PBMC上糖皮質激素受體β的表達,發現糖皮質激素治療不敏感的支氣管哮喘患者糖皮質激素受體β陽性細胞的數量明顯高於糖皮質激素治療敏感的支氣管哮喘患者及正常對照人群,糖皮質激素治療不敏感的支氣管哮喘患者的PBMC在體外培養48 h後,糖皮質激素受體β陽性細胞的數量可以降至正常,但是如果在培養基中同時加入IL - 2和IL - 4,則糖皮質激素受體β陽性細胞的數量不發生變化。而且正常人群的PBMC在體外與IL - 2、IL - 4共同培養後,糖皮質激素受體β陽性細胞的數量也顯著增加。
GR與轉錄活化因子
核因子-κB(NF -κB)是一種分布廣泛、可誘導的轉錄活化因子,對免疫細胞的功能發揮重要的調節作用,NF-κB可以增強許多基因的表達,在哺乳動物的免疫反應及炎症反應中有重要作用,這些基因包括細胞因子、細胞黏附分子、補體和許多免疫受體。
 轉錄
轉錄NF-κB由兩部分組成,即p50蛋白和p65蛋白,NF -κB增強基因轉錄活性的功能可能主要由p65蛋白介導。NF-κB 和糖皮質激素受體在功能上相互抑制,一般來說,兩者之間的物理性相互作用即足以抑制糖皮質激素受體誘導基因轉錄的活性,但糖皮質激素的免疫抑制效應則依賴於p65蛋白和糖皮質激素受體之間更為複雜的相互作用。NF-κB和糖皮質激素受體之間的相互抑制作用最近越來越受到重視,因為這可能在糖皮質激素髮揮抗炎及免疫抑制效應中起重要作用。
McKay 等用NF - κB(p65)、糖皮質激素受體和CAT報告基因共轉染COS -1細胞。當p65的轉染量恆定,而糖皮質激素受體的轉染量增加時,p65介導的NF - κB 應答基因3XMHCCAT的轉錄活性被抑制,並具有量的依賴性。同樣,當糖皮質激素受體的轉染量恆定,而p65的轉染量增加時,地塞米松誘導的糖皮質激素受體增強轉錄的活性也被抑制,並且也有量的依賴性。在培養細胞、人肺組織、人PBMC中存在AP -1和糖皮質激素受體之間的相互作用,這種相互作用對糖皮質激素的抗炎效應有重要影響。Adcock等測定了糖皮質激素治療不敏感的支氣管哮喘患者及糖皮質激素治療敏感的支氣管哮喘患者PBMC上AP -1蛋白與DNA 的結合能力,發現糖皮質激素治療不敏感的支氣管哮喘患者中AP - 1蛋白與DNA的結合力顯著高於糖皮質激素治療敏感的支氣管哮喘患者,與地塞米松共培養60 min後,糖皮質激素治療敏感的支氣管哮喘患者中AP - 1蛋白與DNA的結合力下降5214% ~1112% ,而在糖皮質激素治療不敏感的支氣管哮喘患者中僅下降1515% ~614% (P <0105)
GR介導的轉錄激活作用
 RNA
RNA未與糖皮質激素結合的GR是一個多蛋白的複合體,包括兩分子的HSP90,一分子的HSP70、一分子的HSP56及親免素等。在激素不存在的情況下,這個複合體基本上經歷一個固定循環,解離以及ATP-和HSP-70依賴的重新組合。這種複合體有利於GR蛋白保持在非激活狀態,直到配基激活為止。
糖皮質激素的結合導致的糖皮質激素受體的構象變化現在還未明確,但卻有一系列的功能變化:激素與受體結合在一起導致受體與熱休克蛋白分離,並且不能夠再次結合。此外,部分磷酸化的受體蛋白變為高度磷酸化。最後,在配基結合後,GR蛋白的激素結合區中核內定位的信號將會暴露,並可能導致胞漿蛋白的核轉位。
GR與腫瘤細胞信號傳導
1 GR的激活
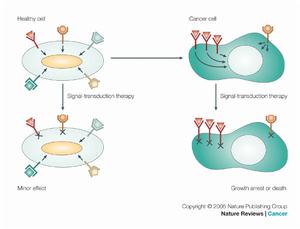 信號傳導
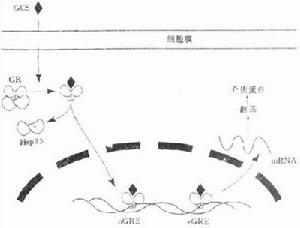
信號傳導GR主要存在於細胞漿中,在未與GC結合前與熱休克蛋白90(HSP90)及小分子親免素等結合成無活性的形式,當GC進入胞漿與GR結合,則HSP90與GR分開,活化的GR二聚化並轉入核內。但GR在腫瘤細胞內的活化可能有異於上述途徑。Catts等證實,將野生型兒童急性淋巴細胞白血病(ALL)細胞暴露於地塞米松或細胞毒製劑中,可導致GR從胞質向核內移位。因而可以推測在ALL細胞中可能有不依賴配體的GR活化。與此相反,Xia等在對人卵巢癌HO-8910細胞的研究中發現,地塞米松可以抑制受胞外信號調節的蛋白激酶1/2的激活,同時使p38的激活大大加快,並且這一作用不能被GR拮抗劑RU486所阻斷。
2 GR與DNA的相互作用
活化的GR以二聚體的形式與靶基因的激素反應元件(hormone response element,HRE)結合,並募集基礎轉錄複合物(BTM)和共激活因子,
 DNA轉錄
DNA轉錄3 GR的調控
GR的調控包括外源性調控和自身調控兩個方面。如使用雌激素激動劑可以下調GR蛋白水平,從而使GR的轉錄活性受到抑制。另一方面,GR還可通過自身調控機制實現信號傳遞途徑之間的對話(cross talking)。當GR與GC結合後,GR mRNA和蛋白質水平均會有不同程度的下調;並且GR與GC複合物同靶基因結合後,還會導致效應細胞的核染色質結構發生變化,使靶基因GRE數目減少,從而下調GR的DNA連線活性影響轉錄激活或抑制。
GR與腫瘤相關基因
激活的GR 可調節多種腫瘤相關基因的轉錄。如Schorr等實驗發現在乳腺上皮細胞中,GC通過GR的介導,不依賴於STAT-5的活性,以劑量依賴的方式誘導了自然bcl-x基因的表達。bcl-x基因作為抗凋亡基因,其高表達提高了乳腺上皮細胞的存活率。與此同時,GR基因本身也受到多種基因和蛋白的調節影響。
 激素書籍
激素書籍Cabral等用人類GR基因啟動子轉染鼠NIH3T3細胞,並檢測到c-jun可抑制啟動子活性的80%,JunB抑制該啟動子活性的30%,而c-fos和JunD對該啟動子無抑制作用。過表達c-jun的NIH3T3細胞克隆顯示了更低的GR mRNA水平。以上結果說明GR基因本身受多種基因調節的影響。
GR 亦可與活化子蛋白-1(actor protein 1,AP-1)和核因子等轉錄因子互動作用,從而調節基因轉錄。Rick等1研究證實,GR和AP-1參與DNA與蛋白複合物的形成,這些複合物在不同乳腺癌細胞系中具有不同的性質和GC誘導性,並可調節癌基因c-fms第一啟動子的轉錄。當AP-1位點發生突變時,會出現該啟動子轉錄活性降低和GC刺激性缺乏。
儘管大量實驗己證實GR與腫瘤相關基因的密切關係,但在腫瘤研究中,仍有許多未知領域等待我們進一步探索。比如已知長期暴露於毒性劑量的砷環境中,可增加患癌風險,Kaltreider等曾通過實驗研究證明無毒劑量的砷可與GR複合物直接作用,並選擇性地抑制GR介導的轉錄,這種轉錄與核功能改變相關,但其具體機制卻不甚明了。
GR與腫瘤細胞凋亡
1 腫瘤細胞的凋亡過程
腫瘤細胞的凋亡是一個複雜的過程,可分為三個階段:起始階段,包括GR介導的基因調節;決定階段,包括生存前和凋亡前因子的抗平衡反應;執行階段,包括caspase和內源性核酸酶的活化。GC誘導的凋亡涉及凋亡的很多方面,如線粒體功能失調和caspase活化,它們在凋亡的所有形式中都是非常重要的步驟。caspase被稱為凋亡執行器,是細胞凋亡調控的關鍵分子群。它們通過切斷與周圍細胞的聯絡,重組細胞骨架,關閉DNA複製和修復,破壞DNA和核結構,誘導凋亡小體的形成等,在細胞凋亡中發揮重要作用。但GC誘導的凋亡與其他形式的凋亡在轉錄水平的起始狀態和多催化功能的蛋白酶和鈣的參與上是不同的。同時,GR與其他傳導通路的交叉連線又增加了GC誘導的凋亡及其調節的複雜性。
 細胞凋亡
細胞凋亡2 GR與凋亡相關基因
GR通過調節凋亡相關基因的表達而誘導或抑制腫瘤細胞凋亡,這些基因主要有bcl-2家族基因,具有誘導增殖和凋亡雙重作用的c-myc基因及腫瘤壞死因子(TNF)等。bcl-2家族基因bcl-x產生兩種不同的蛋白Bcl-x1和Bcl-xs,均可抑制多種類型細胞凋亡,因此有助於腫瘤發生。而與bcl-2同族的bax基因在功能上與其相反,bax表達可加速凋亡。Rogatsky等利用培養的人骨肉瘤細胞作為GR 抗增殖反應的模型,實驗證明在U20S細胞中,活化的GR可引起該細胞發生不可逆的生長抑制、凋亡及bcl-2基因的表達抑制,這種細胞毒反應由GR的轉錄抑制功能介導。c-myc是一種癌基因,在人類多種腫瘤組織中有基因擴增或高表達,當c-myc與bcl-2結合時,可起到協同作用而誘發腫瘤。Moran 等研究發現在永生性人乳腺上皮細胞系MCF10A 的培養基中去除GC,儘管有表皮生長因子和胰島素存在,但仍可誘發凋亡。並發現c-myc的異位表達可促進凋亡,bcl-2的過分表達可抑制凋亡。儘管MCF10A細胞在CD95 (即FAS) 受體激活後可發生凋亡,但去除GC所導致的細胞死亡不依賴於CD95受體的信號通路。TNF是能引起細胞凋亡的主要“死亡因子”。它通過與細胞膜TNF受體結合後,激活細胞自殺程式,引發胞內一系列生化反應,導致凋亡。TNF-a能夠促進細胞內游離鈣濃度增高,激活鈣依賴性核酸內切酶,引起DNA片段和細胞凋亡。
此外,有些化學物質也可以通過GR與GC相互作用的機制,影響某些腫瘤相關基因的表達,參與多種腫瘤細胞凋亡過程。
展望
糖皮質激素(GC)作為一種甾體類激素,
 激素書籍
激素書籍雖然GR的研究仍處在試驗階段,但已看到GR在腫瘤臨床套用中的前景。研究發現GR mRNA兩種剪下變異體GR-α和GR-β的mRNA表達水平與腫瘤細胞GC抵抗的出現有重要關係。GR對某些腫瘤的預後評判也有一定價值,如Ho 等觀察到多數原發性肝癌的胞質中可發現GR,它是原發性肝癌切除術後的獨立預後因素,GR陽性的患者較GR陰性患者生存率低。
綜上所述,GR通過轉錄激活或轉錄抑制調節腫瘤相關基因轉錄和腫瘤細胞凋亡,影響基因產物的合成,從而影響腫瘤細胞代謝的多個環節,相信隨著這方面研究的不斷深入,在不遠的將來,腫瘤發病機制研究及治療方法會有更大的突破。
參考文獻
[1]張方,錢桂生,吳學玲,謝永宏,王興友,陳維中. 人糖皮質激素受體變異體cDNA序列克隆和表達及其活性研究[J]. 第三軍醫大學學報, 2006,(01).
[2]程曉剛. 糖皮質激素受體結構與功能研究進展[J]. 國外醫學.分子生物學分冊, 2003,(01).
[3]崔穎鵬,姜儻. 糖皮質激素受體結構和功能的關係[J]. 國際檢驗醫學雜誌, 2005,(12).
[4]龔德華,王慶文. 糖皮質激素作用機制的新進展[J]. 腎臟病與透析腎移植雜誌, 2000,(01).
[5]矯毓娟,許賢豪. 糖皮質激素受體同型及其與激素敏感性關係的研究進展[J]. 中國神經免疫學和神經病學雜誌, 2006,(03).
[6]黃紅恩,王慶文. 糖皮質激素作用機制研究的新進展[J]. 中國醫學文摘.內科學, 2000,(06).
[7]杜臻雁,唐福林. 糖皮質激素抗炎作用機制的研究進展[J]. 中華醫學雜誌, 2006,(35).
[8]楊珍,湯靜燕. 糖皮質激素受體和激素抵抗機制研究新進展[J]. 國外醫學.兒科學分冊, 2004,(06).
[9]盧建. 糖皮質激素及其受體對細胞數量穩態調節的作用[J]. 基礎醫學與臨床, 2007,(02).
[10]李琳芸,王柳燕,王昌富. 糖皮質激素受體及其檢測方法的研究進展[J]. 微循環學雜誌, 2005,(03).
[11]鍾連生, 馬道銘, 陳志強. 糖皮質激素受體研究進展[J]. 國外醫學.皮膚性病學分冊, 2005,(04)
[12]Chikanza IC. Mechanisms of corticosteroid resistance in rheumatoid arthritis: a putative role for the corticosteroid receptor beta isoform .Ann N Y Acad Sci, 2002, 966 :39-48 .
[13]W ebster JC, Oakley RH, JewellCM, et al. Proinflammatory cytokines regulate human glucocorticoid receptor gene expression and lead to the accumulation of the dom inant negative beta isoform: a mechanism for the generation of glucocorticoid resistance .Proc NatlAcad SciU S A, 2001, 98 :6865-6870 .
[14]Pujols L, Xaubet A, Ram irez J, et al. Expression of glucocorticoid receptors alpha and beta in steroid sensitive and steroid insensitive interstitial lung diseases .Thorax, 2004, 59 :687-693 .
[15]http://baike.baidu.com/view/83722.htm
[16]http://www.hlyyao.com.cn/CYYW011.htm
