
簡介
 屎腸球菌
屎腸球菌屎腸球菌是動物腸道正常菌群中的菌類,因其能產生乳酸,所以歸屬於乳酸菌類。屎腸球菌是一種益生菌,對維持動物腸道菌群生態平衡起到重要作用,特別是在幼齡動物的腸道保健和疾病的防疫和治療上有突出的表現。許多企業將其製成微生態飼料添加劑,但是屎腸球菌的抗逆性較差,在生產加工中不能有效的保持其活性,對屎腸球菌的利用產生了很大的問題。
屎腸球菌,革蘭氏陽性,橢圓形或球桿狀,無芽孢,無鞭毛。生長溫度為30℃~40℃,適宜pH為5.0~7.5。最適生長溫度為35~38℃。屎腸球菌是兼性厭氧的乳酸菌,和大多數的乳酸菌一樣,其耐酸性耐高溫性等抗逆性能差,但是相對於嚴格厭氧、培養和保存條件都苛刻的雙歧桿菌、乳桿菌來說,屎腸球菌更是便於生產和使用的首選菌種。常規的乳酸菌必須在動物腸道的特定部位才能產生功效,天然乳酸菌不耐飼料制粒時的高溫、胃酸及膽鹽的侵蝕。
經飼料制粒後大多數乳酸菌會死亡,在胃裡大約30min後也會死亡殆盡,安全到達腸道發揮作用的活菌數非常低,也無法吸附在腸黏膜的適當位置,隨著糞便排出體外。屎腸球菌代謝可以產生有機酸、過氧化氫細菌素等物質,這些物質具有抑制病原菌和腐敗菌,提高免疫力、改善畜產品品質等生理功效。
其中代謝產生的細菌素可有效治療養殖場中常見的頑固疾病,如對葡萄球菌、梭狀芽孢桿菌、沙門氏菌和志賀氏菌有拮抗作用,並且不留有藥殘。代謝產生大量乳酸,可以顯著降低動物腸道pH,保持腸道的酸性環境,抑制病原菌的生長,對致病菌痢疾桿菌、傷寒桿菌、副傷寒桿菌、彎曲桿菌、葡萄球菌等致病菌有拮抗作用。過氧化氫能抑制和殺死革蘭氏陰性菌和過氧化氫陽性菌如假單胞菌屬、大腸桿菌類、沙門氏菌屬。
耐藥特徵
了解糞腸球菌和屎腸球菌在臨床標本中的分布及對常用抗菌藥物的耐藥性,比較不同標本中兩種腸球菌的耐藥性,為臨床治療提供參考。方法常規法進行菌種鑑定,紙片擴散法進行藥物敏感試驗。結果420株腸球菌屬,來源於尿液、膿液分泌物、痰液,分別占43%、20%、14%位居各類標本中的前3位,糞腸球菌對青黴素、氨苄西林和哌拉西林/他唑巴坦的耐藥率分別為23.0%、17.6%和21.2%,明顯低於屎腸球菌對這3種抗菌藥物的耐藥率(90.0%、87.0%和87.7%),而屎腸球菌對氯黴素和四環素的耐藥率分別為7.9%和24.6%,明顯低於糞腸球菌對這兩種抗菌藥物的耐藥率(43.8%和74.4%),兩種腸球菌中,未發現萬古黴素耐藥株;尿液標本中糞腸球菌對呋喃妥因的耐藥率明顯低於屎腸球菌。結論臨床標本中,腸球菌屬在尿液中的分布最高;糞腸球菌和屎腸球菌的耐藥譜明顯不同,在治療時,應根據不同的菌種選擇用藥,由於腸球菌屬對萬古黴素的耐藥率極低,在重症感染時,可選用萬古黴素進行治療。
耐藥性分析
 屎腸球菌
屎腸球菌一、材料和方法
1、標本來源2004年1月至2007年12月本院住院患者的臨床標本中分離出的144株糞腸球菌和121株屎腸球菌(多次分離的重複株以第一次計算),其中尿80株、痰75株、分泌物27株、血25株、引流液17株、膽汁14株、膿液10株、導管9株、腹水8株。
2、醫院感染的診斷標準按照衛生部醫政司醫院感染監控小組制定的醫院感染的診斷標準。
3、細菌培養鑑定和藥敏試驗按常規的方法對住院患者的各種臨床標本進行細菌的培養分離,用VITEK2全自動微生物鑑定儀及配套試劑進行細菌的鑑定及藥敏試驗;鑑定卡GP、藥敏卡AST-353。
4、質控菌株金黃色葡萄球菌ATCC25923、糞腸球菌ATCC29212。
5、統計學方法WHONET5。4軟體統計彩χ2檢驗,檢驗水準α=0.05。
二、結果
1、HLAR株的分布情況265株中引起醫院感染的HLAR主要分布在ICU占26.8%(71/265),其次腎內科17.0%(45/265)及神經內科8.3%(22/265),感染以泌尿系統感染為主占30.2%(80/265),其次呼吸道28.3%(75/265)、傷口10.2%(27/265)及血液9.4%(25/265)。
2、HLAR株對臨床常用抗生素的耐藥情況265株HLAR腸球菌中糞腸球菌占54.3%(144/265),屎腸球菌占45.7%(121/265),兩者對臨床常用的11種藥物表現為多重耐藥,且屎腸球菌對氨苄西林、阿莫西林/棒酸、亞安培南、環丙沙星、莫西沙星的耐藥率明顯高於糞腸球菌,但糞腸球菌對喹奴普汀/達福普汀的耐藥率明顯高於屎腸球菌(P<0.05),對萬古黴素、替考拉寧均敏感,對力奈唑烷耐藥率分別為0及4.1%。
三、討論
在臨床上由腸球菌引起的感染中以糞腸球菌及屎腸球菌為主,近年來隨著抗菌藥物的廣泛開發與套用,腸球菌對多種抗菌藥物特別是對氨基糖甙類高水平耐藥的增多,使腸球菌成為導致醫院感染的重要致病菌,另由於糞腸球菌與屎腸球菌對抗生素的耐藥有一定的差異,因此為臨床治療腸球菌的感染帶來了挑戰。
從結果可見,對氨基糖甙類高水平耐藥糞腸球菌和屎腸球菌主要引起泌尿生殖道及呼吸系統的感染,分別為30.2%(80/265)、28.3%(75/265),與相關的報導基本相符,糞腸球菌與屎腸球菌的體外藥敏結果顯示其對VAN、TEC均敏感,而對LZD耐藥率為0.0%及4.1%;說明VAN、TEC及LZD對糞腸球菌和屎腸球菌均有較強的體外抗菌活性,提示這三種藥物可作為治療由其引起的重症感染時的經驗用藥。
本次調查結果顯示屎腸球菌對AMP、AMC、IMP等β-內醯胺類的耐藥率均在90.0%以上,明顯高於糞腸球菌的10.0%以下,對CIP、MFX等喹諾酮類抗生素的耐藥率為88.4%、97.5%,明顯高於糞腸球菌的56.30%及47.9%;而糞腸球菌對QD、TE的耐藥率分別為95.8%、78.5%,高於屎腸球菌的41.3%、54.5%,結果與資料報導相近;兩者對紅黴素的耐藥率均大於90.0%,比羅燕萍[3]所報導的偏高。這可能與腸球菌屬內各種之間的耐藥機制不同有關,由此細菌室分離的腸球菌應準確鑑定到種的水平,以便指導臨床合理選用抗生素。
因此,臨床在治療腸球菌引起的感染時一定要注意引起感染的腸球菌的類型而針對性的選用抗生素,同時注意其是否對高水平的慶大黴素或鏈黴素耐藥,因高水平氨基糖甙類耐藥會使得氨基糖甙類與作用於細胞壁的藥物如青黴素類、糖肽類的聯合作用消失。
溶血素形成過程
 屎腸球菌
屎腸球菌其他毒力因子
1、聚集物質AS(aggregationsubstance)
聚集物質是腸球菌的表面蛋白。最初研究表明,它是細菌質粒得以進行接合傳遞的蛋白。以後發現該蛋白除了介導細菌細菌之間的接觸外,也促進細菌與宿主細胞的粘附而與許多感染性疾病相關。研究表明與健康人群來源的菌株相比,臨床來源的菌株聚集物質基因Asa1呈高流行。1998年Schlievert-PM等在研究腸球菌引起的心臟感染的小鼠模型時,發現給小鼠注射含有聚集物質AS及結合物質EBS(enterococcalbindingsubstance,AS受體)的菌株AS+EBS+株,小鼠出現症狀並死亡。給予AS-EBS-株則均存活。進一步在用導管插入術構造心內膜炎的小鼠模型中發現,AS+EBS+菌株產生的贅生物較大,脾腫大更嚴重,認為AS在心內膜炎中起重要作用。
AS聚集物質的致病機制主要是對真核細胞的黏附。聚集物質C端含有LPXTG細胞壁錨著基序結合細菌,N末端含有兩個Arg-Gly-Asp基序即RGD,而相鄰的N末端區可以被巨噬細胞結合,也可被真核細胞表面受體整合素CD11b/CD18(CR3)識別,通過整合素的介導間接提高細菌的黏附能力。體外實驗觀察到AS有助於腸球菌黏附腸上皮細胞、腎小管細胞及心臟內皮細胞。聚集物質還有助於腸球菌粘連至細胞外基質蛋白如:纖維蛋白、纖維連線蛋白、血小板凝血酶敏感蛋白、玻璃粘連蛋白、膠原Ⅰ型等,這種黏附是導致宿主細胞感染的第一步。
2、表面蛋白esp(enterococcussurfaceprotein)
esp基因最初是由shankar在臨床分離的糞腸球菌E.faecalis中發現的,帶有該基因的菌株引起多重感染,後來陸續在屎腸球菌E.faecium中也發現了esp基因。研究表明含有esp基因的臨床分離菌株普遍具有生物膜形成能力如:Esp有助於E.faecalis在膀胱中持續存在,從而提高感染能力。Esp還可促進細菌對無生命物質的黏附並形成生物膜,如體外試驗中esp蛋白可在聚苯乙烯或聚氯乙稀表面形成膜。生物膜的形成可以使細菌的感染更加持久,呈慢性趨勢[65-67]。因此,腸球菌在尿路感染中的分離率較高。
E.faecalis的Esp蛋白是一種由1873個胺基酸組成的細胞壁相關蛋白。esp蛋白與金黃色葡萄球菌的膜形成相關蛋白Bsp結構上具有相似性。其N末端的胺基酸序列(50-743aa)與資料庫的序列無相似性,中間核心區序列(744-1665aa)的球形結構同B型鏈球菌的Calpha及Rib蛋白結構相似,C末端重複序列(1666-1873aa)含有膜形成疏水區及一個較小變異的LPXTGX基序,大多數革蘭陽性菌的胞膜相關蛋白存在此基序。
3、心內膜炎抗原efaA
efaA是一種分泌到血清中結合胞壁的蛋白,是腸球菌的又一表面黏附素蛋白,和口腔鏈球菌的黏附素有顯著的同源性。腸球菌是引起感染性心內膜炎的第三大原因,約導致5-20%自身心內膜炎,6-7%人工心內膜炎病例。腸球菌通過efaA結合心臟組織基質,引起心內膜感染。有報導E.faecalis的efaA在小鼠腹膜炎模型中,也有致病性。
4、膠原結合蛋白ace(accessorycolonizationfactor)
膠原結合蛋白是腸球菌表面的黏附素,又稱粘附聚集因子,屬於MSCRAMM家族(microbialsurfacecomponentsrecognizingadhesivematrixmole-cules),可以介導腸球菌黏附膠原I型和膠原IV型及層粘連蛋白等,是腸球菌的毒力因子[70]。在不同類腸球菌中有不同的名稱,在糞腸球菌中為Ace,在屎腸球菌中稱Acm。Ace在糞腸球菌E.faecalis中檢出率比較低。屎腸球菌Acm與金黃色葡萄球菌膠原黏附素Cna總體有62%的同源性,也是潛在的致病因子。
5、明膠酶gelE(gelatinase)
據報導幾乎一半的臨床分離菌株中有明膠酶表達,而在健康個體中明膠酶分離率約為27%。明膠酶在E.faecalis和E.faecium臨床菌株中均可分離到。腸球菌感染的動物模型表明:明膠酶及其調節基因fsr(enterococcusfaecalisregulator)具有致病作用,序列分析表明:腸球菌的明膠酶與銅綠假單胞菌的胰蛋白酶(33KDa)以及桿菌的中性蛋白酶相似。實驗表明:明膠酶可以水解明膠、膠原、酪蛋白、血紅蛋白和其它生物活性肽,引起細菌擴散,參與炎症的進程。
6、胞外的超氧化物O-2
很多E.faecalis及一些E.faecium等菌株,可以產生超氧化物,而且其產量比一般的非致病菌株多。如Huyke-MM等人報告臨床分離的標本中有95%的E.faecalis,38.5%的E.faecium產生胞外O2-。氧自由基可以破壞細胞膜,使腸球菌通過薄弱的上皮屏障進入血液。超氧化物還可以引起紅細胞溶解,並可使組織損傷更嚴重。
7、透明質酸酶
透明質酸酶是細菌表面相關的酶,能促進細菌在結締組織的傳播。腸球菌也能分泌透明質酸酶。Rosan和Williams在研究致齒周圍疾病的微生物時發現腸球菌透明質酸酶和化膿性鏈球菌的透明質酸酶有60%的相似。透明質酸酶也可能是腸球菌的致病因子,在侵襲性疾病中可能起作用。
8、脂磷壁酸(lipoteichoicacid)
它是革蘭染色陽性菌細胞壁上普遍存在的由親水的多聚磷酸甘油骨架通過酯鍵與疏水的糖脂結合形成的兩性分子聚合體,在許多革蘭染色陽性菌中,它是炎症因子的刺激物。Ehrenfeld等報導純化的來源於E.faecalis的脂胞壁酸抑制由性信息素誘導的菌細胞的聚集。其致病作用可能與調節炎症反應和促進質粒轉移有關。
9、性信息素(sexpheromone)
性信息素是E.faecalis分泌的一個小的線形肽段(7-8胺基酸),誘導腸球菌的asa1等基因,表達聚集物質,促進菌株之間質粒的接合轉移,及粘附宿主細胞。同時性信息素還具有人白細胞趨化劑的作用,引起粒酶的釋放和呼吸爆發。所以,性信息素也是腸球菌的致病因子。腸球菌的AS基因,存在於不同質粒上,由不同的性信息素誘導產生相應的聚集物質。性信息素的編碼基因有cpd、ccf、cob、cad等。
10、磷脂酶
磷脂酶在其它細菌中是毒力因子,但作為腸球菌的毒力因子,目前缺少足夠的證據,只在E.faecalis有35%的分離率,在E.faecium中的分離率為4%。
上述致病因子中各毒力基因並不同時出現,研究還發現上述致病因子基因型和表型並不一致,雖然腸球菌致病相關因子的發現使人們對於腸球菌的致病性有著更加明確的認識,但其中的機制尚未徹底明了,並且有些因子的確認還不具備充分的證據,如磷脂酶、透明質酸酶、超氧化物等。因此需要進一步從分子水平進行研究,為臨床的治療提供理論支持。
生物學特性
 屎腸球菌
屎腸球菌1、屎腸球菌對熱的耐受性
益生菌是對動物生長和生產都有很大幫助的菌類,可以調節胃腸道的菌群平衡,使優勢菌占優勢,調節動物腸道生理功能,提高動物的免疫力,提高飼料的利用率。在飼料加工的過程中,有益菌能否耐受制粒時的溫度將是檢驗有益菌是否有經濟價值的重要因素。大多乳酸菌在飼料加工的過程中死亡殆盡,或者活性降低很多。李梅等(2007)從健康仔豬糞便中分離出的屎腸球菌能耐受45℃的溫度,在50℃情況下幾乎不能存活。Risley在研究飼料制粒對微生物添加劑的影響時發現,嗜酸乳桿菌不能耐受制粒,60℃即檢不到活菌,酵母不能耐受80℃的制粒溫度。優良的益生菌的選擇,溫度是至關重要的方面,在屎腸球菌菌株的篩選中選取耐高溫的菌株或者改變微生態製劑的形態來避免高溫帶來的損害在現在的科學條件下應該是可行的。
2、屎腸球菌對胃酸的耐受性
豬胃環境中pH一般在3左右,屎腸球菌適宜pH為5.0~7.5,通過篩選可以選出耐胃酸的菌株。NelsonPérezGuerra等(2007)試驗中所用的屎腸球菌(CECT410)在模擬胃液pH2.0中能存活;李梅等(2007)從40日齡健康仔豬空腸粘液中分離到的兩株屎腸球菌(Ef1、Ef2),在pH4.0顯陽性,但在pH3.5顯弱陽性;沈中艷等(2007)從健康仔豬新鮮糞便中篩選分離到的屎腸球菌能耐受pH2.5合成胃液達3h,且在pH1.5合成胃液培養30min後有存活,活菌數降低2~5個數量級,但培養達3h後無存活;陳平潔等(2001)從4月齡健康豬糞中成功分離出兩株屎腸球菌(SF1、SF2),在pH2.5條件下存活率僅為0.004%,在pH3.0條件下可存活1.4%。屎腸球菌對不同胃酸pH耐受能力的不同,原因有很種,比如動物品種與年齡、分離的部位、及篩選的方法不同。對於相同動物來源的的屎腸球菌對胃酸有不同的耐受程度,很大程度上是與篩選的方法不一樣造成的。
使用的方法是從40日齡空場黏膜為材料進行腸球菌的分離的,將屎腸球菌分離後再對其生物學特性進行鑑定;沈中艷等(2007)是選擇仔豬的新鮮糞便為材料篩選腸球菌的,先將材料在特定的培養基上培養後,篩選出乳酸菌,然後再此基礎上進行生物學特性上的篩選試驗,選擇活性好耐特定病原菌較強的菌株,最後鑑別為屎腸球菌。此方法有很強的目的性,篩選出的屎腸球菌是特異型菌株。其篩選出的屎腸球菌的耐酸性很強可達pH1.5。而陳平潔等(2001)選用4月齡豬的內層糞便做為材料進行篩選的,再進行屎腸球菌的分析鑑定。屎腸球菌的耐酸性試驗結果的不同的原因取決於他們對菌株的來源的選擇和篩選的方法。初生仔豬胃內pH一般在5~6之間,出生後由於高產酸細菌定植而逐漸下降,數小時後pH可降到4,然後緩慢下降,在2月齡前pH可保持在3左右,而成年豬胃酸pH會更低。要想使屎腸球菌作為飼料添加劑使用,必須要過胃酸這一關,否則無法使屎腸球菌發揮出正常的生理作用。
3、屎腸球菌對膽鹽的耐受性
乳酸菌進入腸道後,小腸中膽鹽的高滲透壓環境不利於其存活,屎腸球菌對膽鹽的耐受性相對來說較好,李梅等(2007)分離出的屎腸球菌可在豬膽鹽40%條件下顯弱陽性。沈中艷等(2007)分離出的屎腸球菌可以耐受3%的膽鹽濃度。哺乳仔豬膽囊內膽汁分泌量很少,膽汁量在出生後3周內緩慢增加,小腸內膽鹽濃度在0.03%~0.30%範圍內波動。大量試驗表明,篩選得到的屎腸球菌對膽鹽有良好的耐受性。對膽汁的敏感性是篩選益生素菌株的重要指標。美國Altech生物技術中心規定用於益生素的細菌對膽鹽應有70%以上的抵抗力。可見,屎腸球菌具有良好的耐膽鹽性能及對胃腸道逆性環境的耐受性,具備了在小腸定植的基本條件。
4、屎腸球菌在腸道的定植能力
益生菌能夠在胃腸道上皮細胞上粘附並生長繁殖時其發揮作用的基本條件之一,是保證有效的競爭排斥病原菌的定植,並發揮其生物功能的前提,益生菌在腸上皮細胞上粘附力的強弱也是篩選益生菌菌株的重要標準之一。Tuomola等(1998)研究了12種不同乳桿菌的粘附情況,結果發現乾酪乳桿菌、嗜酸乳桿菌、乾酪乳桿菌鼠李糖亞種(LC-705)和乳桿菌GG(ATCC53103)的粘附能力最強。而屎腸球菌的粘附力僅次乾酪乳桿菌。乳酸菌能順利通過胃腸不利環境,在腸壁上的粘附是其定植並大量繁殖變成優勢種群的前提。
5、屎腸球菌對抗生素的耐受性
腸球菌,尤其是屎腸球菌具有對廣譜抗生素的耐藥性(BRUNTON,1984;Murray,1990;Woodford等,1995;Gold等,1996;Rice等,1996;Klare等,1997;Quintiliani等,1999;Kak等,2002;Klare等,2002)。陳平傑等(2001),屎腸球菌(SF1、SF2)對飼料添加劑中大多數常用抗生素都具有耐藥性,但在液態下進行的耐藥性試驗中對萬古黴素、四環素、亞氨硫黴素等少數幾種抗生素較為敏感,但是屎腸球菌在飼料中是以固態形式存在的,兩者的接觸面積不如液態狀態下大,藥敏性抗生素對屎腸球菌的殺滅作用有限。來源於食品的屎腸球菌對氨苄西林、環丙沙星、慶大黴素和鏈黴素敏感,其原因可能是屎腸球菌的來源不同,其耐藥性能也不同。
6、屎腸球菌的抑菌作用
除了對抗生素具有耐藥性外,還對常見病原菌、強毒菌株有較強的抑制作用,Jin等(2000)研究證明腸球菌能夠抑制大腸桿菌K88ac和K88MB粘附到小腸黏膜上,並對K88ac的抑制作用具有劑量效應,當活菌數達到109時抑制效率在90%以上。這可能是屎腸球菌代謝過程中產生乳酸、乙酸、異丁酸、乙醇、2,3-丁二醇、過氧化氫、細菌素等物質,這些物質具有抑制病原菌和腐敗菌的作用。其中代謝產生的細菌素對葡萄球菌、梭狀芽孢桿菌、沙門氏菌和志賀氏菌等有拮抗作用。細菌素是一類選擇性作用於靶細胞的抗菌物質,對特異性病原菌有很好的殺滅作用,如豬鏈球菌病在我國為一種常見的豬病,在我國其病原菌主要以馬鏈球菌獸疫亞種為主。該病在豬群中以呼吸道為主要傳播途徑,且可以通過傷口、消化道等途徑傳播,發病率和死亡率較高。目前全群預防在臨床上常用弱毒疫苗免疫和飼餵抗生素兩種方法。由於致病性豬鏈球菌的血清型較多,因此免疫預防常常不能引起很好的效果。抗生素雖能夠在一定程度上控制該病的流行,但容易引起豬胃腸道菌群失調,造成二重感染和內源性感染及肉製品藥物殘留等問題,用抑制病原菌的細菌素來治療將是一個極佳的選擇。
對抗生素耐藥性分析
 屎腸球菌分析
屎腸球菌分析腸球菌屬是醫院感染的主要條件致病菌之一,可引起人體各種器官的感染。隨著抗菌藥物的廣泛套用,腸球菌屬耐藥性有所增加,特別是耐高濃度氨基糖苷類抗生素及耐萬古黴素腸球菌的增多,腸球菌引起的醫院感染有明顯增多的趨勢。為了解腸球菌的臨床感染及其耐藥特性,對近年臨床分離的腸球菌及其體外藥敏情況進行了分析。
腸球菌可引起尿路感染、菌血症、肺炎、腦膜炎、心內膜炎等多種感染症。本實驗結果顯示,在近3年多時間中,共分離到383株腸球菌,主要為糞腸球菌和屎腸球菌占87%。菌種來源顯示,尿液占首位(50.9%),其次為膿液分泌物(225%),痰標本占第三位(18%),以上3類標本共占91.4%。腸球菌所致感染最多見於尿路感染,多與尿路器械的操作、流置導管、尿路結構異常有關,為僅次於大腸埃希菌的第二位引起尿路感染的病原菌。創傷及肺部感染也較為常見,可能與創傷暴露部位的感染以及介入性的操作(呼吸機、插管)等容易並發腸球菌感染有關。提示應加強此類疾病的醫院內感染的預防和控制。
研究顯示,萬古黴素和力奈唑烷對3種腸球菌均保持較高的敏感性(耐藥率<10%),腸球菌對四環素耐藥率為40%-55%之間,對青黴素的耐藥率>80%。糞腸球菌對呋喃妥因的耐藥率9.2%,對莫西沙星、左鏇氧氟沙星耐藥率分別為36.2%、44%。屎腸球菌對呋喃妥因的耐藥率為48.8%,對莫西沙星和左鏇氧氟沙星的耐藥率分別為82.7%、86.6%。高濃度慶大毒素和高濃度鏈黴素的耐藥率在不同菌種間差異較大,糞腸球菌分別為36.2%、33.8%,而屎腸球菌則為63%、58.2%,鶉雞腸球菌為39.4%、54.5%,這些耐高濃度慶大黴素和高濃度鏈黴素的菌株,是因為產生了氨基糖苷鈍化酶,當它們與作用於細胞壁的抗菌藥物聯用時不具協同作用,從而給臨床抗感染治療帶來困難。屎腸球菌與糞腸球菌及鶉雞腸球菌進行比較,屎腸球菌的耐藥率明顯偏高,提示屎腸球菌的耐藥機制可能與糞腸球菌及鶉雞腸球菌不同,因此,臨床實驗室應對腸球菌鑑定到種的水平,以便指導臨床合理使用抗生素。
耐力古黴素腸球菌(VRE)的出現給臨床治療帶來了困難,其耐藥機制主要為細胞壁肽聚糖前體末端發生改變,使萬古黴素不能與之結合而失去抗菌活性。VRE所致嚴重感染目前尚無滿意可靠的治療方法,應依據藥敏試驗結果和臨床情況選擇用藥,並宜聯合用藥。許多文獻報導,利奈唑胺在體外對糞腸球菌及屎腸球菌包括VRE均有較好的抑菌活性,並經美國FDA批准可用於治療耐萬古黴素屎腸球菌感染。
近年來,國內外報導耐萬古黴素的腸球菌呈逐漸增加的趨勢,這就提示臨床合理使用萬古黴素,以保持腸球菌對萬古黴素的高度敏感性。
套用研究
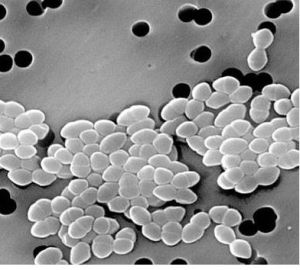 屎腸球菌
屎腸球菌1、屎腸球菌對仔豬生產性能的影響
有試驗表明在仔豬日糧中添加屎腸球菌(CECT410)顯著提高斷奶後28d仔豬的體重(P<0.05)。英國利茲大學(UniversityofLeeds)初步研究表明,在日糧中添加屎腸球菌能改善斷奶後一周仔豬的生產性能。Maeng等(1989)認為,在小豬日糧中一直添加屎腸球菌(SF68)到4個月,能增加小豬體重、採食量及提高飼料轉化率。張振斌等(2004)試驗表明,腸球菌組顯著提高斷奶後兩周仔豬的採食量,有提高仔豬日增重的趨勢。但Broom等(2006)試驗表明,在仔豬日糧中添加屎腸球菌對斷奶仔豬的生產性能沒有顯著影響,這與Taras等(2007)結果一致但是屎腸球菌能使飼料轉化率有所改善。
2、屎腸球菌對斷奶仔豬腹瀉率的影響
Taras等(2007)試驗證明腸球菌可減少大腸桿菌的量,進而減少仔豬腹瀉的發生率(P<0.05),這與他以前所報導的結果相同(Taras等,2006)。Nelson等(2007)試驗證實在斷奶仔豬日糧中添加篩選具有一定抗逆性的四種潛在乳酸菌(包括腸球菌)可以增加仔豬採食量,提高日增重及飼料轉化率,並可以顯著的減少糞便中大腸桿菌的數量(p<0.05),降低腹瀉率。禹慧明等(2000)試驗表明,含腸球菌的活菌製劑用於0~28日齡仔豬,可使腹瀉次數減少35.6%。張振斌等(2004)試驗表明,在斷奶後兩周內腸球菌製劑有降低仔豬腹瀉率的趨勢。Foulquié-Moreno等(2006)報導腸球菌(E.faeciumSF68)用作益生素防治抗生素相關性腹瀉,在臨床上認為是有效的。
3、對仔豬免疫功能的影響
腸道菌群不僅影響散布於消化道淋巴組織(GALT)中免疫細胞的機能,還可以影響全身的免疫系統(Travnicek等,1989)。還有一些研究表明,不同菌群效應可激活免疫系統。另外,腸道菌群對營養抗原類物質的免疫耐受具有誘導和維持作用。益生菌在改善動物健康和防禦病原體方面發揮了重要的作用,如雙歧桿菌和其它乳酸菌可以增加腸道的IgA(Takahashi等,1998;Vitini等,2000);乾酪乳桿菌具有免疫佐劑活性(Perdigon等,1991)及植物乳桿菌促進感染大腸桿菌的小鼠抗體的產生(Herias等,1999)。ViolaStrompfová等(2006)試驗表明,添加屎腸球菌(EK13)分別在7d和14d後極顯著或顯著提高新生仔豬血清總蛋白濃度(P<0.01;P<0.05);能降低血清膽固醇(P<0.01;P<0.05);能提高血紅蛋白、血細胞比值和白細胞數量(P<0.01;P<0.01;P<0.05);能提高紅細胞數量(P<0.01)、粒性白細胞的吞噬活性指數(P<0.01)、血鈣水平(P<0.01)及血清谷胱甘肽過氧化物酶濃度(P<0.05)。也有報導表明,在日糧中添加屎腸球菌(SF68)對斷奶後6d仔豬的血清IgG及腸內IgG無影響,但有降低斷奶後20d仔豬血清IgG濃度的趨勢(P<0.01)。細胞因子是宿主全身免疫系統的重要調節者。由於散布在消化道淋巴組織(GALT)上抗原(益生素)與淋巴樣細胞的相互作用,促使細胞因子的釋放,這些細胞因子能調節機體全身的免疫系統。因此,添加屎腸球菌(SF68)有降低仔豬血清IgG的趨勢,這就提示了屎腸球菌可作為一種免疫調節劑。
4、對腸道菌群及腸黏膜結構的影響
Strompfova等(2009)試驗證明屎腸球菌EK13可以顯著降低仔豬糞便中大腸桿菌的數量。NelsonPérezGuerra等(2007)試驗發現,添加屎腸球菌(CECT410)顯著降低斷奶仔豬糞便中大腸桿菌的活菌數量(P<0.05),這與ViolaStrompfová等(2006)結果一致。說明在日糧中添加屎腸球菌能有效促進有益菌的增殖,減少有害細菌的數量,從而改善腸道內環境。也有報導表明,添加屎腸球菌(SF68)對斷奶後6d仔豬小腸內厭氧菌或乳酸菌無影響,這與Scharek等(2005)結果相似,出現的不同研究結果可能與所添加的屎腸球菌的菌株不同,其到達腸道中發揮作用的數量不同造成的。大量研究表明,採食水平高低與仔豬小腸黏膜絨毛高度及隱窩深度有關,高採食水平可促進小腸黏膜的發育(Pluske等,1996;VanBeersSchreurs等,1998;McCracken等,1999)。張振斌等(2004)試驗發現,添加腸球菌製劑能改善斷奶後兩周末仔豬小腸黏膜絨毛高度及絨毛高度與隱窩深度比值,且斷奶後兩周內仔豬採食量顯著增加;因此,外源添加屎腸球菌通過提高仔豬採食量促進小腸黏膜的發育,從而增進對營養物質的消化吸收功能。
5、屎腸球菌對肥育豬母豬的影響
肥育豬和母豬胃酸一般較仔豬pH低,約在1~3,鑒於屎腸球菌的耐酸範圍,在pH小於3的情況下,通過胃後活菌的數量會減少很多,存活下來的活菌數量不是很多。針對於包被型的屎腸球菌,其受胃酸的影響較小,通過胃後活菌的數量會較多,因此在使用上沒有太大的影響。到目前國內外鮮有對屎腸球菌在肥育豬和母豬上的使用報導,屎腸球菌代謝過程中產生乳酸的量較多,可以在肥育豬上使用,測其生長性能及飼料轉化效率。在母豬上的使用,可以減少糞便中大腸桿菌和沙門氏菌的含量,在保證母豬體內微生態平衡的情況下,保持保育舍中的微生態平衡,減少環境中有害菌的含量,保證乳豬生活在一有害菌少的環境中,減少攝入體內有害菌的量。鑒於屎腸球菌的良好的抑菌性能,同時可以減少哺乳母豬乳房炎及其它炎症的發生率。
潛在危害性
腸球菌感染是新生兒敗血症的第3位病因,近幾年新生兒和兒童腸球菌敗血症的發病率增加了6倍。據報導腸球菌是內源性和外源性醫院感染的第二大病原菌,檢出率僅次於大腸桿菌。有資料統計,在引起尿路感染的致病菌中,腸球菌感染居第2位;腹腔、盆腔感染,腸球菌居第3位;敗血症,腸球菌居第3位,病死率12.6%~57%。當抗生素大量使用或宿主免疫力低下時,宿主與腸球菌之間的共生狀態失衡,腸球菌離開正常寄居部位進入其他組織器官,它首先在宿主組織局部聚集達到閾值密度,然後在粘附素的作用下粘附於宿主細胞的胞外矩陣蛋白,分泌細胞溶解素、明膠酶等毒力因子侵襲破壞宿主組織細胞,並通過質粒接合轉移使致病性在腸球菌種間擴散,並耐受宿主的非特異性免疫應答,引起感染性疾病的發生髮展。
同時,腸球菌還極易形成耐藥性,其耐藥性包括固有耐藥、獲得性耐藥及耐受性3種。由於其細胞壁堅厚,腸球菌對許多抗生素表現為固有耐藥。腸球菌的耐藥性在20世紀70年代表現為對氨基糖甙類耐藥,如慶大黴素和鏈黴素,80年代表現為耐β-內醯胺類及糖肽類,1986年首次發現耐萬古黴素腸球菌(VRE)。90年代以後,由於抗菌藥物廣泛套用,加之侵入性治療以及過度使用氟喹諾酮類和口服頭孢菌素類藥物等因素,導致腸球菌耐藥菌所致感染菌株及病例不斷增加,已成為院內感染的主要病因。由於這類細菌所引起的耐藥菌感染治療難度大,加之腸球菌對抗生索的耐藥現象比較複雜,目前,在臨床分離的腸球菌中有許多是多重耐藥菌株。
美國醫院內監測系統表明由VRE導致的醫院感染1989年為0.3%,1993年為7.9%,在ICU內則增加至13%,至1997年,超過15%的醫院內腸球菌感染為VRE。VRE耐藥譜廣、易造成感染和流行並且能將耐萬古黴素基因轉移給其它革蘭氏陽性菌,如金黃色葡萄球菌等。同時腸球菌屬在乾燥的物體表面可存活數日至數周,無疑可以從被污染的手和衣物表面進行攜帶傳播。腸球菌屬具備毒力和流行性,本身又可產生介導炎症反應的能力,以及它的耐藥性和播散性(包括耐藥因子的播散),都對臨床患者有潛在的危害,加重病情的進展。
另外,美國研究人員在《醫學微生物學雜誌》上發表文章指出,糞腸球菌能產生有害化學物質,破壞DNA,進而引起促發直腸癌的基因活動。他們在實驗室中仔細研究了直腸細胞對糞腸球菌的反應。結果發現在發酵狀態下,糞腸球菌會生成一種叫“超氧化物”的氧分子,這種氧分子會破壞周圍細胞的DNA,但糞腸球菌的影響遠不止於此。這種超氧化物會給被稱為巨噬細胞的免疫細胞發出強烈信號,改變一些腸道寄生細胞的生長、分裂方式,甚至還會增強致癌基因的活動。研究證實,發酵狀態下,與人體重要生理過程相關的42種細胞基因都受到了糞腸球菌的影響。
因此,InternationalProbioticsAssociation和世界衛生組織(WHO)均建議:益生菌中不宜使用腸球菌!目前最新的益生菌產品如東海藥業生產的阿泰寧、寶樂安、爽舒寶等都不含有腸球菌,預計在不久的將來,我國政府也會出台限制腸球菌用於微生態製劑的規定。
