
簡介
 翼龍
翼龍翼龍是生活在晚三疊世(lateTriassic)至白堊紀(Cretaceous)時期的一種能在空中飛行的爬行類動物,屬於恐龍的近親和鄰居。也是最早出現的飛行的脊椎動物。1801年居維葉將這類飛行爬行動物命名為翼手龍(pterodactyl)。其特徵是前肢第四指骨加長加粗,支撐由身體側面延展的皮膜,形成翅膀。三疊紀晚期出現,白堊紀末滅絕。全球分布。
分類
在歷史上,翼龍目的科學分類是很困難的,因為它們的化石紀錄有許多間斷,許多新發現填補了這些間斷。傳統上,翼龍目被分為兩個亞目:
喙嘴翼龍亞目(Plieninger,1901):一群早期、原始翼龍類,許多物種有長尾巴,翼上有短指骨。它們體型小,手指仍然適合攀爬。它們出現在晚三迭紀,持續到晚侏羅紀。喙嘴翼龍類是並系群,翼手龍類從它們演化而來,而非兩者有共同祖先;所以隨者親緣分支分類法研究的增加,本名詞逐漸很少在科學文獻中出現。
翼手龍亞目(Plieninger,1901):更多樣性的翼龍類,有短尾巴,翼上有長指骨。它們出現在中侏羅紀,在白堊紀-第三紀滅絕事件中滅絕。
發現歷史
 翼龍化石
翼龍化石第一個翼龍化石是在1784年由義大利自然學家Cosimo Collini發現。Collini當時將這些動物誤認為海生動物,將它們的長前肢充當槳來使用。一些科學家持續支持這個海生動物假設,直到1830年,德國動物學家Johann Georg Wagler仍提出翼手龍屬的前肢是用來游泳的。在1809年,喬治·居維葉(Georges Cuvier)將一個在德國發現的物種命名為翼手龍屬(Ptero-dactyle),並首次提出這種動物是種飛行動物。然而因為科學名稱的標準化,這物種的正式屬名改成Pterodactylus。
自從1784年在索倫霍芬石灰岩層發現第一個翼龍類化石後,當地沉積層已發現29種翼龍類。在1828年,瑪麗·安寧(Mary Anning)在英國萊姆里吉斯發現著名的雙型齒翼龍。大多數翼龍類化石不是保存良好。它們的骨頭是中空的,當沉積物堆積在它們身上時,骨頭會被壓平。保存最良好的化石在1974年發現於巴西的Araripe Plateau。當沉積物堆積到這些化石上時,沉積物會壓縮骨頭,而非壓碎骨頭。
大多數古生物學家認為翼龍類是采動力飛行,而非原先認為的滑翔。翼龍類化石已在北美、南美、英國、歐洲、非洲、亞洲、澳洲等地發現,除了南極洲以外。已發現至少60屬翼龍類,體型小如小型鳥類,大的翼展可達10公尺長。
體態結構
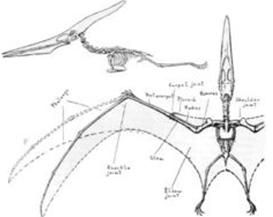 骨骼重建圖
骨骼重建圖 翼龍類的生理結構因為飛行演化的需求,而與它們祖先的生理結構有大幅改變。翼龍類的骨頭中空,內有空氣,類似鳥類的骨頭。它們擁有胸骨,可讓飛行肌肉附著在上,還有加大的腦部,顯示出與飛行有關的特化特徵。
翼
翼龍類的翼由皮膜與其它軟組織構成,由不同形式的緊密纖維補強。皮膜連線極長的第四手指與身體側面之間。連線到腕部的翅骨(Pteroid),是一種翼龍類專有的骨頭,協助支撐腕部到肩膀的前膜(Propatagium)。翅骨可能可以往前擺動延展前膜,但論點這是非常有爭議的。在有些較晚的翼龍類里,肩膀的脊骨固定至Notarium(一種在鳥類與某些翼龍類肩膀出現的複合脊骨),可在飛行時使身體堅挺,並提供肩胛骨穩定支撐。
古生物學家們經常爭論翼膜是否也連線到後肢。喙嘴翼龍類的索德斯龍、以及蛙嘴龍科的熱河翼龍、還有桑塔納岩層發現的翼手龍類化石,證實翼膜的確連線至後肢,至少某些物種有連線。然而,現代蝙蝠與飛鼠翼膜的連線方式上有相當大的不同,而不同種類的翼龍類可能也有不同的翼膜連線方式。研究顯示翼龍類的四肢比例有相當大的不同,可能反映了不同的翼展方式。許多翼龍類有蹼狀腳掌,可能不是全部都有,蹼狀腳掌可能具有氣動上的作用,但也有論點認為蹼狀腳掌是種游泳的證據。某些現代滑翔動物也具有蹼狀腳掌,例如鼯猴。
毛
翼龍類並沒有羽毛證據,但它們的獨特處在於它們覆蓋者毛,類似哺乳類的毛,但非同源演化。翼龍類的毛並非哺乳類的真實毛髮,而是獨特的結構,為趨同演化的後果。雖然在有些例子裡,翼膜上的纖維被誤認為毛,有些化石的頭部與身體上的確有毛的印痕,例如索德斯龍,毛不類似現今蝙蝠的毛,這是另一個趨同演化的例子。毛的出現是基於飛行的需求,也顯示翼龍類是溫血動物。
神經系統
一個使用X光線掃描翼龍類腦部的研究,顯示出它們習性的訊息。研究翼龍類頭顱骨是非常困難的,因為它們頭顱骨非常易碎,但俄亥俄大學的Lawrence Witmer與他的同僚使用X光計算機斷層掃描,建立起兩個物種的腦部3D影像(明氏喙嘴翼龍、桑塔納古魔翼龍)。 發現這兩個物種小腦有巨大的絨球(Flocculus)。絨球是用來整合來自關節、肌肉、皮膚、與平衡器官的訊息。
翼龍類的絨球占了它們腦部體積的7.5%,比其它脊椎動物還多。鳥類的絨球也比其它動物大,但只占腦部的1%~2%。
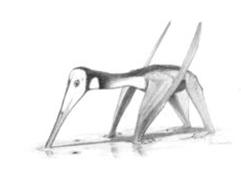 行走想像圖
行走想像圖絨球傳送神經訊號,讓眼部肌肉產生小而自動的運動。這讓動物的視網膜產生穩定的影像。翼龍類有大型絨球的原因,可能是因為它們巨大的翼,巨大的翼意味者有大量的感應訊息要處理。
地上運動
翼龍類的臀窩是稍微往側上方,股骨頭適度的往內側彎,顯示翼龍類是半直立步態。當它們飛行時,大腿可能抬高到與身體平行。
曾有爭論翼龍類在地上移動時,是四足還是二足。顯在已發現大量的翼龍類足跡,足跡上有明顯的後肢四趾與前肢三趾;已證實翼龍類的確以四足在地上移動。有論點指出較小的翼龍類有較長的後肢,例如雙型齒翼龍,除了飛行以外,行走與奔跑時可能采二足方式。其它小型翼龍類,例如喙嘴翼龍,奔跑時使用四肢。較大的翼龍類有較小的後肢與大型身體前半部,一般認為它們在地面上移動時使用四肢。根據所發現的翼龍類足跡化石,可以發現它們正在涉水或找食物,還沒有發現可證明它們在飛行或滑翔的足跡化石。
演化與滅絕
起源
 頭顱骨化石
頭顱骨化石 翼龍類的骨骼結構因為適應飛行而有重大改變,而且沒有它們最直接祖先的描述,所以對翼龍目祖先了解不多。因為它們的踝部結構,翼龍類被認為與恐龍是近親。已有數個相關理論,其中近年最盛行的是類似Scleromochlus的鳥頸類初龍,或者是類似沙洛維龍的原蜥形目動物。
因為翼龍類被證實沒有樹棲生活的演化適應,所以它們的飛行演化途徑被認為跟鳥類不同路線,鳥類的飛行演化途徑是「從樹往下的」。大多數方案認為翼龍類從長腿的陸地奔跑動物演化而來,如Scleromochlus或沙洛維龍,上述兩者都有皮膜,從後腿延展至身體或尾巴。這些研究顯示翼龍類的飛行演化途徑是“從地面往上”,或是攀爬懸崖。
在2008年,一份研究顯示最早的翼龍類是群樹棲、食蟲的小型動物。
滅絕
一般認為早期鳥類的競爭,導致許多翼龍類滅絕。到了白堊紀末期,只有發現大型翼龍類。較小的物種滅絕,並由鳥類取代。 白堊紀末滅絕事件滅絕了恐龍、翼龍類、與許多其它動物。其它人提出大多數翼龍類往依靠海洋的生活發展。所以當這次滅絕事件嚴重地影響翼龍類賴以為生的海洋生物時,翼龍類滅絕了。化石紀錄中缺乏小型翼龍類,可能是因為鳥類競爭,或是因為它們的骨骸難以保存下來。
發現過程
世界上第一枚翼龍胚胎化石,在中國遼西熱河生物群被古生物學家汪筱林、周忠和博士發現。相關研究論文刊登在世界最著名的學術刊物———英國的《自然》雜誌上。由於這樣一塊無比珍貴化石的發現,獲得強有力的證據,證明翼龍這類與恐龍同時出翼龍(2張)
現又同時絕滅,比鳥類早約7000萬年飛向藍天的中生代空中霸主不是胎生,而是像其他爬行動物和鳥類一樣卵生。
中國科學院古脊椎動物與古人類研究所的研究員汪筱林一項重大發現是一個尚未出殼的翼龍的化石,發育中的翼龍胚胎,骨骼了了,清晰可辨。化石產自遼西錦州義縣一個湖泊沉積的地層中,距今約1.21億年。這一地點曾經發現大量保存精美的狼鰭魚、滿洲鱷以及一些翼龍和鳥類化石。
胚胎化石傳達了最明確的信號:翼龍,與鳥類和其他爬行動物一樣是卵生而非胎生。汪筱林介紹,翼龍是恐龍的近親,二者生活在同一時代,是第一個飛向藍天的爬行動物,人們經常將翼龍誤認為是“會飛的恐龍”。翼龍起源於約2.2億年前的晚三疊世,絕滅於6500萬年前的白堊紀末期。當恐龍占據著陸地時,翼龍卻一直控制著天空。
1784年義大利的古生物學家科利尼在德國發現第一件翼龍化石時,不能確定它屬於哪一類動物,有人認為它生活在海洋中,也有人認為它是鳥和蝙蝠的過渡類型。
1801年,法國著名的比較解剖學家居維葉鑑定它為翼龍,歸於爬行動物。翼龍化石的發現比恐龍早了半個多世紀,自從翼龍化石被發現,人們對這類非常奇特的最早飛向天空的動物就充滿了好奇,一直在探求。
種群演化
起源
生存於三疊紀晚期的斯克列羅龍,可能是翼龍類祖先的近親,翼龍類的骨骼結構因為適應飛行有大幅改變、特化,而且沒有它們最直接祖先的化石證據,所以對龍目的演化起源了解不多。因為它們的踝部結構,翼龍類被認為與恐龍是近親。已有數個相關理論,其中最盛行的理論是類似斯克列羅龍(Scleromochlus)的鳥頸類主龍、能以二足方式站立的原始主龍形類派克鱷或者是類似沙洛維龍的原蜥形目動物。翼龍類專家大衛·安文認為這些動物因為個別的生理特徵,都不符合翼龍類祖先的假設。翼龍(2張)
克里斯·班尼特、大衛·彼得斯曾先後提出翼龍類屬於原蜥形目,或者是原蜥形目的近親。大衛·彼得斯利用電腦繪圖軟體,企圖找出原蜥形目、翼龍目的關聯。克里斯·班尼特在試圖找尋翼龍類、恐龍的趨同演化特徵時,發現翼龍類除去後肢特徵時,相當類似原蜥形目,提出原蜥形目、翼龍類有演化關係。
2007年,麥可·班頓(MichaelBenton)等人提出不同看法,認為翼龍類與恐龍是近親,即使不考慮後肢特徵,翼龍類仍非常類似恐龍。班頓等人認為雖然目前沒有翼龍類最直系祖先的化石證據,但翼龍類很明顯是屬於主龍類,尤其是鳥頸類主龍演化支。根據他們的分析,翼龍類是斯克列羅龍的姊妹分類單元,或者是從介於斯克列羅龍與兔鱷之間的物種演化而來。
2008年,研究顯示最早的翼龍類是群樹棲、食蟲的小型動物。
2011年,斯特林·奈斯比的早期主龍類研究也支持翼龍類、斯克列羅龍互為姊妹分類單元的理論。因為翼龍類被證實沒有樹棲生活的演化適應,所以它們的飛行演化途徑被認為跟鳥類不同路線,鳥類的飛行演化途徑是“從樹往下的”。大多數方案認為翼龍類從長腿的陸地奔跑動物演化而來,如斯克列羅龍或沙洛維龍,上述兩者都有皮膜,從後腿延展至身體或尾巴。研究顯示翼龍類的飛行演化途徑是“從地面往上”,或是攀爬懸崖。
鼎盛時期
繁盛於中生代侏羅紀(2.08億年前至1.44億年前)和白堊紀(1.44億年前至6,640萬年前)的飛行爬蟲類的成員。翼龍目(Pterosauria)屬於爬蟲類的古龍亞綱(Archosauria)。恐龍類和鱷類也屬於古龍亞綱,鳥類是古龍類的後裔。三疊紀(2.45億年前至2.08億年前)的古龍類趨向兩足步態,前肢自由地作其他方式的套用。鳥類和翼龍類二者同樣將前肢轉變成翼。滅絕原因
傳統理論認為,早期鳥類的競爭,導致許多翼龍類滅絕。到了白堊紀末期,只有發現大型翼龍類;而小型翼龍類已消失,生態位被早期鳥類取代。但是,化石紀錄缺乏小型翼龍的現象,也可能是因為它們的骨架脆弱、難以保存所致,而跟鳥類的生物多樣性無關。白堊紀末滅絕事件滅絕了恐龍、翼龍類、與許多其他動物。另一種解釋是,大多數翼龍類發展成依靠海洋的生活模式。所以當白堊紀末滅絕事件嚴重地影響翼龍類賴以為生的海洋動物時,翼龍類跟者滅絕。但是,神龍翼龍科與帆翼龍科是棲息於內陸,並非依靠海洋維生。馬斯特里赫特階,例如:鳥掌翼龍科、無齒翼龍科、夜翼龍科、古神翼龍科。這些新發現物種,顯示白堊紀晚期的翼龍類仍保持多樣性,相對於白堊紀早期
史前大型動物
| 生物的進化是一個很複雜的過程。但生物的進化無不是經歷由簡單到複雜、由水生到陸生、由低等到高等這樣一個漫長的演化過程。而今,這些動物都不復存在了,因為它們的後代在自然條件的影響下經常發生變異,適應自然條件的動物可以生存、發展,而不適於自然條件的動物則被淘汰,這種適者生存的過程就叫做自然選擇。 |
