
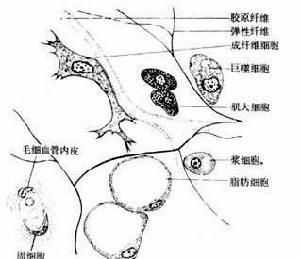
 細胞間識別
細胞間識別類型
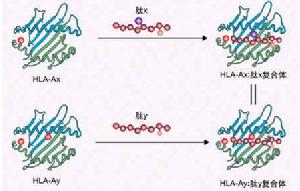 植物細胞間識別
植物細胞間識別植物細胞間識別
在植物中,細胞間識別可以涉及兩個不同物種的細胞,如根瘤菌和豆科根細胞;也可以發生在同一物種的細胞之間,如花粉粒和柱頭。細胞識別依賴細胞表面的多糖、糖蛋白和糖脂等分子中的特有順序。一般認為負責識別表面糖順序的是植物凝集素。花粉粒和柱頭之間相互識別就是植物凝集素作用的一個實例,甘藍科識別系統的分子組成是由S基因複合體編碼的,分子組成包括柱頭表面的一個大分子糖蛋白和花粉粒表面一個能識別它的植物凝集素。識別促使柱頭釋放水分,花粉粒吸水、伸出花粉管、生長並和卵受精。但是,花粉粒只在表達S基因複合體的不同等位基因的植物中才能引起上述一系列反應。如果花粉粒先經過從表達相同等位基因的柱頭提純的糖蛋白處理,它就不能在一個正常可配伍的柱頭上發芽。這就說明了識別的專一性。
動物細胞間識別
 動物細胞間識別
動物細胞間識別 激素識別就屬這一類。激素通過血流影響分布全身各部分的靶細胞,以協調各種生理活動。許多激素是蛋白質,也有一些是甾體。甾體激素不溶於水,但可與專一的載體蛋白結合而成為可溶性的;一旦從載體釋放,它們就通過靶細胞的細胞膜,和細胞質中專一的甾體激素受體蛋白結合,然後到細胞核中進一步起作用。每一甾體激素只識別一個受體蛋白,但是,同一受體蛋白在不同的靶細胞中可調控不同的基因,也就是說,受體是相同的,被激活的基因卻不相同。因此,同一激素和相同的受體蛋白結合,在不同細胞中所引起的效應也就不同。
另一種信號分子和激素不同,它們非常迅速地被附近靶細胞攝取,或者,就在合成部位附近被專一的酶破壞,以致進入血循環的量一般是微不足道的。如分屬9類的16個不同的前列腺素中,有許多都能和不同的細胞表面受體結合併具有不同的生物效應。
複雜性
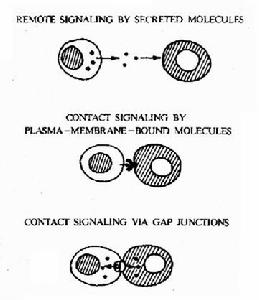 細胞間識別--神經系統
細胞間識別--神經系統在胚胎髮育中,識別的現象也起重要作用。已經從海膽精子的頭部(頂體)分離出結合蛋白,這種分子只和同種卵的質膜結合;另一方面,也發現卵子質膜中含有物種專一的糖蛋白,它和結合蛋白在粘著中相互作用。哺乳類中則是精子質膜中含有一種物質,可以和卵子透明帶中的專一糖蛋白直接結合以達到受精。
視網膜和頂蓋神經原之間的識別具有嚴密的區域性。從視網膜背部長出的軸索必然到達對側頂蓋的腹部,而從腹部長出的必然到達對側頂蓋的背部。在實驗條件下,如果將視網膜切成背腹兩半,分別將細胞分散,用同位素標記背部細胞,然後再與腹部細胞混和;頂蓋也切成背腹兩半,分別與視網膜細胞一起培養,觀察同位素標記細胞分別聚集在頂蓋背腹兩部分數量。結果發現視網膜背部的細胞大多聚集在頂蓋的腹部,而腹部的細胞大多集中在頂蓋的背部。這說明視網膜細胞與頂蓋相當部位的細胞之間存在著相互識別的能力。
動物中另一種細胞識別的形式是相同細胞間的相互粘著。由於在整體中觀察研究細胞間的相互粘著比較困難,常用的方法是使某一組織分散為單個細胞,再觀察它們的重新聚合。用這種分散聚合的方法,在海綿和胚胎組織都分別取得了有意義的結果。
信號分子
 細胞間識別--信號分子
細胞間識別--信號分子從化學結構來看細胞信號分子包括:短肽、蛋白質、氣體分子(NO、CO)以及胺基酸、核苷酸、脂類和膽固醇衍生物等等,其共同特點是:①特異性,只能與特定的受體結合;②高效性,幾個分子即可發生明顯的生物學效應,這一特性有賴於細胞的信號逐級放大系統;③可被滅活,完成信息傳遞後可被降解或修飾而失去活性,保證信息傳遞的完整性和細胞免於疲勞。
從產生和作用方式來看可分為內分泌激素、神經遞質、局部化學介導因子和氣體分子等四類。
從溶解性來看又可分為脂溶性和水溶性兩類。脂溶性信號分子,如甾類激素和甲狀腺素,可直接穿膜進入靶細胞,與胞內受體結合形成激素-受體複合物,調節基因表達。水溶性信號分子,如神經遞質、細胞因子和水溶性激素,不能穿過靶細胞膜,只能與膜受體結合,經信號轉換機制,通過胞內信使(如cAMP)或激活膜受體的激酶活性(如受體酪氨酸激酶),引起細胞的應答反應。所以這類信號分子又稱為第一信使(primary messenger),而cAMP這樣的胞內信號分子被稱為第二信使(secondary messenger)。目前公認的第二信使有cAMP、cGMP、三磷酸肌醇(IP3)和二醯基甘油(DG),Ca2+被稱為第三信使是因為其釋放有賴於第二信使。第二信使的作用是對胞外信號起轉換和放大的作用。
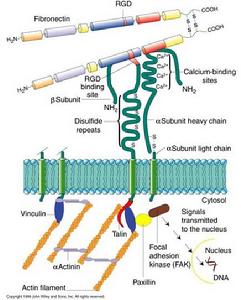
受體
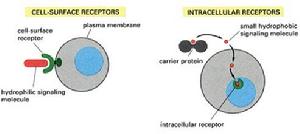 細胞間識別--受體
細胞間識別--受體根據靶細胞上受體存在的部位,可將受體分為細胞內受體(intracellular receptor)和細胞表面受體(cell surface receptor,圖8-2)。細胞內受體介導親脂性信號分子的信息傳遞,如胞內的甾體類激素受體。細胞表面受體介導親水性信號分子的信息傳遞,可分為:①離子通道型受體、②G蛋白耦聯型受體、③酶耦聯型受體。
每一種細胞都有其獨特的受體和信號轉導系統,細胞對信號的反應不僅取決於其受體的特異性,而且與細胞的固有特徵有關。有時相同的信號可產生不同的效應,如Ach可引起骨骼肌收縮、降低心肌收縮頻率,引起唾腺細胞分泌。有時不同信號產生相同的效應,如腎上腺素、胰高血糖素,都能促進肝糖原降解而升高血糖。
細胞持續處於信號分子刺激下的時候,細胞通過多種途徑使受體鈍化,產生適應。如:
①修飾或改變受體,如磷酸化,使受體與下游蛋白隔離,即受體失活(receptor inactivation)。
②暫時將受體移到細胞內部,即受體隱蔽(receptor sequestration)
③通過內吞作用,將受體轉移到溶酶體中降解,即受體下行調節(receptor down-regulation)
蛋白激酶
 蛋白激酶
蛋白激酶1、內分泌(endocrine):內分泌細胞分泌的激素隨血液循環輸至全身,作用於靶細胞。其特點是:①低濃度,僅為10-8-10-12M;②全身性,隨血液流經全身,但只能與特定的受體結合而發揮作用;③長時效,激素產生後經過漫長的運送過程才起作用,而且血流中微量的激素就足以維持長久的作用。
2、旁分泌(paracrine):細胞分泌的信號分子通過擴散作用於鄰近的細胞。包括:①各類細胞因子;②氣體信號分子(如:NO)
3、突觸信號發放:神經遞質(如乙醯膽鹼)由突觸前膜釋放,經突觸間隙擴散到突觸後膜,作用於特定的靶細胞。
4、自分泌(autocrine):與上述三類不同的是,信號發放細胞和靶細胞為同類或同一細胞,常見於癌變細胞。如:大腸癌細胞可自分泌產生胃泌素,介導調節c-myc、c-fos和ras p21等癌基因表達,從而促進癌細胞的增殖。
作用
 細胞間識別-- 變形體聚集
細胞間識別-- 變形體聚集變形體之所以聚集,是因為在飢餓的情況下,變形體開始合成並分泌環腺苷酸(cAMP)。由於還沒有弄清楚的原因,某些細胞成為聚集中心,所分泌的cAMP和其他飢餓的變形體表面的專一受體結合。每一cAMP脈衝向四周擴散,誘發它周圍的個體移向cAMP來源的方向,並分泌它們本身的cAMP。這個新的cAMP脈衝決定移行的方向,並誘發它周圍的細胞分泌cAMP。如此,從每一個聚集中心引發的cAMP以搏動的方式傳播,以同心圓或者螺鏇形擴展到更大的範圍,引起大量個體的聚集。自由生活的單細胞變形體就這樣聚集成多細胞蛞蝓體。
在飢餓的情況下,變形體不但分泌cAMP並對它發生反應,同時它們開始相互粘著。有一些和細胞粘著有關的分子只有飢餓的變形體才合成,用免疫的方法已經證實它們是細胞表面糖蛋白。
細胞表面糖蛋白和糖脂的低聚糖的複雜性,及它們在細胞表面的外露位置,都提示它們可能在細胞識別中起重要作用。有的細胞具有和專一的低聚糖結合的表面蛋白。這樣的相互作用已在植物細胞中得到證明,在動物細胞雖然也有不少間接的事實,但是,究竟細胞表面的這類分子是否也在細胞識別中起同樣的作用還難以斷定。
