
海洋生態
正文
━━━━━━━━━━━━━━━━━━━━━━━━海洋環境
海底地形
海洋的水平分區
海洋的垂直分層
海洋緯度自然帶
海流
海水的化學組成
常量元素
營養元素
微量元素
溶解氣體
有機物
海底沉積
海洋生物
浮游生物
游泳生物
底棲生物
底上生物
底內生物
底游生物
潮間帶生物
海洋生態系統
海洋食物鏈
生產者
初級消費者
次級消費者
分解者
能流
深海生物群落
━━━━━━━━━━━━━━━━━━━━━━━━
海洋環境
地球上海洋面積共3.6億平方公里,約占地球總面積的71%,為陸地面積的2.4倍。海洋之間並無隔斷,由於海水的水平和垂直運動,還不斷地進行著水體和物質的混合和交換。海洋平均深度約3820米。目前已知海洋最深處超過11000米。通常水深每增加10米,就增加1個大氣壓。因此海底最深處的壓力可達1100個大氣壓。
海底地形 海底構造複雜,有海底山脈、高原、盆地、峽谷和海溝等各種地形。通常將海域分為:①大陸架,海洋靠近陸地的部分,從沿岸起緩慢往外傾斜,平均坡度僅0°07′,即向外延伸1公里,平均加深1.5米左右。大陸架的寬度與陸地地形有關,有些海洋幾乎沒有大陸架,而中國沿海大陸架寬度有幾百公里。大陸架水深可達200米。②大陸坡,在大陸架以外,其主要特點是坡度明顯增大(有的可達20°~30°),水深也急劇增加,平均可達2440米左右。③大洋盆地,在大陸坡以外,地形廣闊而平坦,是海洋的主要部分,水深可達6000米左右。④海溝,狹而長,水深超過6000米以上,最深的海溝是馬里亞納海溝,水深達11022米。
海洋的水平分區 與大陸架相對應的淺海稱為近岸區或近海區,大陸坡以外的廣大海域稱為大洋區。
近海區面積小,僅占海洋總面積的8%,但由於陸地大量營養物質注入,因此是生物生長、繁殖的重要場所。
海洋的垂直分層 大洋區的水層可垂直劃分為若干帶。①上層帶,下限為浮游植物的補償深度,即光合作用產生的氧恰與呼吸作用消耗的氧相等的深度,其具體數值取決於光強和水的透明度,一般10~120米,平均約為50米。②中層帶,起於上層帶的下限,下至200~300米深處。③次中層帶,水深可達600~700米。④半深帶,水深由600~700米至2000~2500米,幾乎無光。⑤深層帶,水深2500~6000米,無光。⑥深淵帶,水深在6000米以下,無光。
海洋水溫通常由表層向下遞減,到達一定深度,溫度降低顯著,即溫躍層。深層帶和深淵帶水溫一般是1~4℃。許多海洋生物往往只能在一定水層生活,如浮游植物生活在上層帶。
海洋緯度自然帶 海洋表層水溫在一年中的變化,隨水體所處緯度而定。按照表層水溫可把海洋劃分為若干自然帶。①熱帶,表層水溫最低在20℃以上,溫度變化範圍小,一般僅2~3℃。②亞熱帶,最低水溫在15℃以上,溫度變化範圍較大,可達6~7℃以上。③溫帶,水溫在15℃以下,可近於0℃,溫度變化範圍最大。④近極區,水溫最高不超過10℃,溫度變化範圍不超過2~3℃。⑤極區,水溫最高不超過0℃(或5℃),溫度變化範圍很小,沿岸常有冰層覆蓋。
海流 對海洋環境起著積極主導的作用。海流包括潮流、風海流、坡度流、升降流(上升流和下降流)、密度流、渦流和大洋環流等。大洋環流是大洋海流的骨幹,而上升流區域則是重要的漁場。
太平洋黑潮暖流經中國台灣海峽的東面,沿東海邊緣向東北方向流去。它的兩支分流對中國沿海有明顯的影響。一支分流經巴士海峽流入南海;另一支分流經東海注入黃海,在夏季甚至可達渤海。這兩支海流的方向大體都是由南向北,是暖性的。中國沿海另一支海流是中國沿岸流,北起黃海和渤海,沿海岸南下,經台灣海峽注入南海。沿岸流受大陸氣候影響,冬夏溫差大,沿途容納了大量淡水,所以鹽度也較低。在台灣以東海域、舟山附近、海南島東北部、粵東和閩浙沿海有上升流。除海流外,波浪、潮汐、光、溫度和水壓力等都是影響海洋生物的重要物理因素。
海水的化學組成 已發現海水含有80多種元素,其化合物的種類則更多。這些物質以離子態(陽離子、陰離子、絡合離子)、懸浮態、膠體以及氣泡等形式存在於海水中,海水總的含鹽量在35‰左右,但在河口可低至1~2‰。個別水蒸發量大的海域含鹽量偏高,如紅海為40‰。按組分含量的不同,可把海水中的化學成分分為5類:
常量元素 如鈉、鎂、鈣、鉀、鍶、氯、硫、碳、溴、氟、硼等,占海水中溶解鹽類的99.8~99.9%,它們在海水中的含量基本不變。
營養元素 包括氮(以硝酸鹽、亞硝酸鹽和銨等形式存在)、矽、磷、鐵、錳、銅等,這些元素與生物的生長關係很密切。
微量元素 種類很多,但總量卻非常少,僅占總鹽量的0.1%左右,如鋰、銣、銫、鋅、鈾等。
溶解氣體 主要來自海氣交換,如氮、氧、氬、二氧化碳、氖、氦等,在缺氧情況下,還會出現硫化氫、甲烷等。
有機物 包括碳水化合物、脂肪、蛋白質和元素有機化合物等等。部分來源於大陸河流輸入,大部分來自海洋生物的分泌、排泄和生物死體的分解。
海水的鹽度、pH值、含氧量、二氧化碳含量、營養鹽和有機物以及一些微量元素的含量,對海洋生物都有直接的影響。
海底沉積 海底的地形、結構以及沉積物的組成、理化特性(如粒度大小、有機質含量、氧化還原電位等)是影響底棲生物生活的重要環境元素。海底沉積物來源大致是:陸源性沉積、陸上或海底火山的噴出物、無機性沉澱或宇宙產物,以及由生物死體所造成的細泥。在淺海區域,底部沉積物的組成主要受陸地物質來源的影響,海流對沉積物的轉移起主導作用。絕大部分深海或大洋底層的沉積物,都是由海洋生物的殼和骨骼所造成,在大洋的底部,生物屍體的沉積有的竟厚達數千米以上。在近海區,沉積速率一般較高;而在大洋區,沉積速率很慢,每千年僅沉積1厘米左右。
海洋生物
海洋中幾乎到處都有生物,但不同的環境,生物群落的種類組成和結構,以及各種群數量、個體大小、形態、生理生化特性等都很不同。海洋生物分布的格局是與海洋環境相互作用、協調進化的。一般常按生活方式將海洋生物分為浮游生物、游泳動物和底棲生物3類。
浮游生物 無游泳能力或游泳能力弱,懸浮於水中隨水流移動,包括細菌、浮游植物(如硅藻、甲藻等)和浮遊動物(如水母、腹足綱軟體動物的翼足類、異足類,許多海洋動物的幼蟲等)。多數終生營浮游生活,稱為永久性浮游生物;少數種類僅於生活史的某個階段營浮游生活,稱為階段性浮游生物,如許多海洋動物的幼蟲;也有些原非浮游生物,被水流沖盪而出現在浮游生物中,如某些低等甲殼類的介形類、漣蟲類等,稱為暫時性浮游生物。浮游植物只能生活在有光的水層;浮遊動物則不然,有的可以生活在千米以下的深水中,而且多數能在水中作垂直移動。浮游生物一般體重輕(外殼重量輕、體內脂肪含量高,富有粘液)、沉降阻力大(身體相對面積大,體表多刺毛、突起,群體連成片)或者具有纖毛、鞭毛而有一定的運動能力。
游泳生物 指那些具有發達的運動器官而游泳能力強的動物。包括魚類、哺乳動物(如鯨、海豚、海豹)、爬行動物(如海蛇、海龜)、軟體動物(如烏賊、章魚)和一些大型蝦類(如對蝦、龍蝦)等,它們的游泳能力、速度和方式也有很大差異。有些種類能橫跨大洋作長距離的洄游,如鮪魚等;有的游泳很快,如劍魚每小時速度達70公里以上。海鰻以整個身體彎曲擺動向前遊動,烏賊和章魚則從漏斗口向外噴射水流以推動身體反向運動。它們有時生活在上層水域(如太平洋鯡魚),有的生活在中層或底層水域(如小黃魚、真鯛、牙鮃)。游泳動物除了有發達的游泳器官外,身體大多呈流線型,以減小阻力,提高游泳速度。
底棲生物 生活在海洋水域底部和不能長時間在水中遊動的各種生物,包括底棲植物(幾乎全部大型藻類和紅樹等種子植物),底棲動物(海綿、腔腸、環節、線形、軟體、甲殼、棘皮、脊椎等門類均有底棲種)。底棲生物按其與底質的關係,又可區分為底上、底內和底游3大生活類型;在岸邊還存在潮間帶生物。
底上生物 生活於硬質海底。包括在各種底質上營固著生活(如海葵、水螅、藤壺、牡蠣、柄海鞘)、附著生活(如紫貽貝、附著硅藻、大型藻類)以及匍匐海底(如馬糞海膽、刺參和紅螺等)的動物。固著生活的動物運動器官退化,但觸覺器官發達,被動取食,幼體營浮游生活,靠海流擴大其分布區域。固著或附著在船底、浮標、工廠排污管道、海上石油平台和海上其他設施的生物被稱為污損生物(包括微生物、藻類和動物)。它們對交通、軍事和生產均有危害。
底內生物 生活於軟質海底。有的分泌粘性物質形成管子或用分泌物粘結沉積顆粒成管子而棲身其中(如巢沙蠶、磷沙蠶等);有的具有發達的挖掘器官,將身體埋於泥沙中(如黃島長吻柱頭蟲、中國蛤蜊、文昌魚等);有的鑽木(如船蛆)、鑿石(如波紋溝海筍)。
底游生物 有一定的運動能力,能在水底緩慢地行動(如梭子蟹)。
潮間帶生物 一類特殊類型的底棲生物。經過長期的進化,底棲生物對潮汐的漲落、海浪的衝擊、陽光的暴曬、溫度的變化等有很好的適應能力。由於潮間帶生態位的差異,生物在潮間帶的分布很有規則,群落結構比較清楚(圖1)。
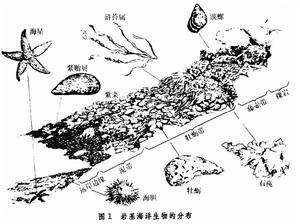 海洋生態
海洋生態海流和水團決定了浮游生物群落的種類組成。根據對南黃海和東海浮游生物的調查,發現群落結構與這一海域海流和水團的性質密切相關,大致可分為:①南黃海中部群落,分布區域與黃海冷水(低溫、高鹽)分布範圍相吻合;②沿海群落,分布在沿岸30海里以內範圍,受中國沿岸流(廣溫、低鹽)控制;③東海外海群落,處於台灣暖流和黑潮暖流流經的區域(高溫、高鹽)。每個群落均有代表種,且一年四季又有所變動。
海底沉積物的特性對底棲生物群落有明顯影響。據調查(1963),中國青島膠州灣底棲動物的區系完全符合與地形、海底沉積及淤泥程度相依的規律,可分為5個明顯的生物群落(表1)。
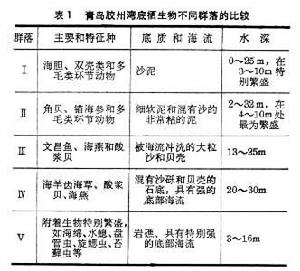 海洋生態
海洋生態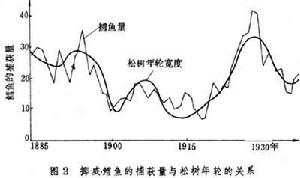 海洋生態
海洋生態海洋生態系統
生態系統的最基本特徵是,能量通過食物鏈不斷地單向流動,物質在食物鏈與無機環境之間不斷地反覆循環。(圖3)表示出海洋生態系統的結構。
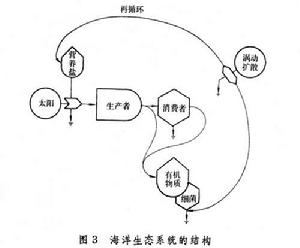 海洋生態
海洋生態食物鏈的長短不一,長的可有5~6個環節,雜食性動物還可同時占據不同的營養級。
海洋動物對食物的選擇不如陸地動物嚴格,能攝食多種食物。在海洋中尚未發現在陸地上見到的那種單食性種類。例如,中國閩南-台灣淺灘漁場的二長棘鯛是底棲魚,其食料生物種類廣泛,包括長尾類(細鰲蝦、褐蝦)、底棲端足類(鉤蝦亞目)、瓣鰓類(短齒蛤、鳥蛤、簾蛤)、介形類(尖尾海螢)和多毛類(沙蠶、似柱蟲),還攝食櫻蝦類(日本毛蝦)、糠蝦類、輻蛇尾、海膽類、星蟲類、腹足類、掘足類、短尾類、海葵類和魚類。海洋動物的捕食與被捕食關係相當複雜。大多數動物的幼體和成體的食性往往不同,可以處於不同的營養級。鯡魚的幼體為箭蟲所攝食,而箭蟲卻是成體鯡魚的一種重要餌料。
生產者 海洋中,能行光合作用的生物有5類:大型定生藻、海洋被子植物、浮游植物、底棲硅藻和光合細菌。儘管在近岸海域大型定生藻的初級生產也很可觀,但就整個海洋初級生產力來說,個體很小的浮游植物是主要生產者,這與淡水環境近似而與陸地環境有很大差別。小型浮游植物所製造的有機物約占海洋初級生產力的95%。因此,海洋初級生產力調查,主要是測定海洋浮游植物的光合作用速率。
影響海洋初級生產力的因素主要是光、海況、營養鹽、浮游植物的種類、生物量和被捕食情況等。
海洋生態系統的能量輸入可分為兩大類,一類來自太陽的輻射,另一類來自物理環境。運動引起的能量被稱為代謝外能量。只有太陽輻射能高、代謝外能量輸入也高的海域,才能夠得到高的生產力,如近岸水域和上升流區域(圖4)。
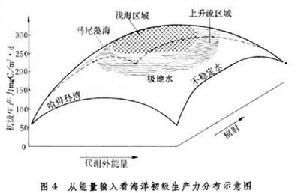 海洋生態
海洋生態光是海洋初級生產力的一個重要限制因素。若已知光隨水深的衰減率、太陽輻射的量以及水中浮游植物色素的數量,即可按以下公式:
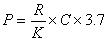
有人曾用此公式計算阿拉斯加灣的初級生產力,所得結果是1.34克(有機碳)/(米2·天),這與實測初級生產力1.50克(有機碳)/(米2·天)很接近。
浮游植物的生長離不開光,但並非所有的浮游植物都能適應強光。因此浮游植物的峰值大多出現在其光強相當於入射光能量大約 1/3的水深處。有些能夠移動的浮游植物,如甲藻,還能根據光強調節所處水深。不同種類的浮游植物對光的利用率有差異,硅藻還藉助輔助色素提高對光的利用率。
海洋初級生產力除了有地域差異時,還有日變化和季節變化。在北緯50°以北海域,初級生產力的高峰通常出現在中午。而在亞熱帶和熱帶海域,高峰出現在中午以前。青島膠州灣浮游植物有明顯的季節變化,在2月和8月,浮游藻細胞總量呈現出高峰,而6~7月和9月則是低谷期。
海流和水團對初級生產力有明顯的影響。在西北太平洋亞北極水域和混合區水團的初級生產力高於亞熱帶水團和黑潮流域。
浮游植物種類的組成和比例是水域初級生產力高低的重要標誌。硅藻占優勢的水域,生產力也高。青島膠州灣及附近水域,浮游植物種群結構以硅藻和甲藻兩大類為主,尤其是硅藻,無論在種數或細胞個數,都占絕對優勢。灣內硅藻的細胞數量可占浮游藻細胞總量的99.9(1979.2)~96.0%(1997.7)。
影響海洋初級生產力第二個重要因子是營養鹽。其中氮和磷在浮游植物生長中起最主要限制作用,在某些水域束毛藻能直接從水中吸收分子氮,因而磷成為限制因子。相反,在某些沿岸水域,由於有大量含磷廢水輸入,氮又可能成為限制因子。一般情況下,氮磷比大約是15~16,比率在10~20範圍內均能滿足生長的需要。在溫帶淺海水域,春季浮游植物的大量繁殖,主要與冬季水混合時將大量營養鹽輸入真光帶有關。上升流區域初級生產力高,主要是大量營養鹽從海底帶上升到水錶層,使硅藻等浮游藻大量繁殖所致。根據營養鹽含量和初級生產力高低,水域被分為富營養、寡營養和中等營養水域。
除了氮、磷外,矽是硅藻生長的一個重要因子,鐵和一些溶于海水中的有機物,如螯合劑、維生素等,通過對浮游植物生長、繁殖的促進或抑制,也直接對初級生產力產生影響。
在沿海水域,大型海藻、底棲微型藻、紅樹林、海草等的生產力也值得重視。這是因為它們的生產量也相當可觀。據估計西太平洋地區海藻的潛在年生產量可達3.0×106噸(鮮重),全球底棲微型藻可達109噸(碳)。不少海洋動物直接以它們為餌料,比如鮑魚直接吃海帶類海藻,有些蝦類直接以紅樹林碎屑為餌料。
初級消費者 植食動物多是濾食性種類,以浮游植物或有機顆粒為食。主要包括橈足類等小型甲殼動物、被囊動物、毛顎動物、水母等浮遊動物,貽貝、牡蠣、扇貝等底棲動物,以及植食性魚類等。浮遊動物與浮游植物雜居在同一水層,在水中能作垂直移動,其種群組成和數量也有明顯的季節變化。
海洋動物的生產力,指一定時間內一群動物所增加的身體物質總重量。但浮遊動物生產力測定有相當困難,常只能間接估算。
動物的生產效率,通常用P/B表示,其中P表示生產量,B表示某一種群的生物量。根據現有的資料,海洋無脊椎動物的生產效率大致如表2。
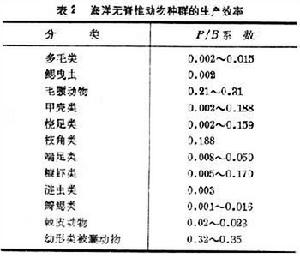 海洋生態
海洋生態次級生產力與個體大小、發育階段、餌料質量和數量、攝食率、同化效率、溫度等有直接關係。有的動物濾食率很高,比如一個海鱒類個體,每分鐘可濾食100毫升,它對餌料沒有選擇性。海洋植食性動物的同化效率比陸地動物高,可達80~90%。
除攝食外,有些無脊椎動物以及海洋動物幼蟲,還能直接從水環境中吸收溶解的有機物質(胺基酸、脂肪酸、多醣及其水解產物)。無脊椎動物幼蟲從溶解有機物質直接吸收的碳可能占其代謝需要量的21~57%。曾有報導,海島哲水蚤對無機和有機顆粒有識別能力,能從混合物中選擇攝食有機物。
次級消費者 即肉食動物,以植食性動物或其他肉食性動物為餌料。因此,不同肉食性動物在食物鏈上可以處於不同的營養級。例如,北方河口的動物,依其營養關係可分為植食性動物、雜食性動物、低級肉食性動物、中級肉食性動物和高級肉食性動物 5類。據調查(1981),閩南-台灣淺灘漁場66種經濟魚類中,低級肉食性動物主要攝食植食性動物和雜食性動物,其種類和數量最多,共有42種,如金色小沙丁魚、鮐魚、二長棘鯛、銀黃姑魚、青石斑魚、日本竹莢魚等。中級肉食性動物主要攝食低級肉食性動物以及植食性和雜食性動物,其種類和數量比較少,共有16種,如大黃魚、中國團扇鰩等。高級肉食性動物主要攝食低級和中級肉食性動物,以及雜食性動物,其種類和數量最少,只有8種,如帶魚、日本馬鮫、路氏雙髻鯊、沙拉真鯊等。
有些肉食性動物的攝食量很高。比如生活於南極海洋的藍鯨,每餐可攝食1噸的磷蝦。海洋動物的食性廣泛,不僅在生長的不同時期採食種類不同,而且隨著季節的不同,食物的組成也有差異。
通常魚獲量與浮游生物的生物量呈正相關。在沿岸,每年7~9月有上升流,在此期間水域的浮游植物和浮遊動物的生物量呈現峰值,而沙丁魚也出現最大捕獲量(圖5)。
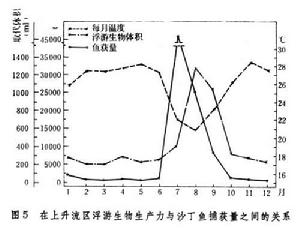 海洋生態
海洋生態海洋動物排出的糞便(糞粒、糞球)含有未消化的有機殘渣,沉入海底後,成為某些底棲動物的餌料。
沿著海洋食物鏈營養級位而上,生物個體也逐漸增大。捕食者與捕獲物的比例,按重量比是100:1,按長度比是4.6:1。鯨是個特例,它是現今地球上最大的動物(體重可達百噸),但卻以個體僅幾厘米的磷蝦為餌料。
食物鏈的長短不一,以微型浮游植物為初級生產者的大洋水域,食物鏈長些,可達5~6個環節,如下左:
 海洋生態
海洋生態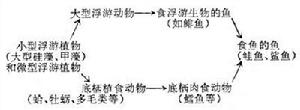 海洋生態
海洋生態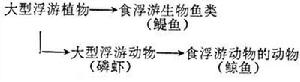 海洋生態
海洋生態以大型浮游植物為主要初級生產者的上升水域的食物鏈大多很短,如上右:
分解者 主要是異養的微生物,它們借分解海洋動植物的死體和其他有機物質獲得能量,同時把有機物逐漸降解還原為無機物。海洋中的碳循環、氮循環、磷循環等與陸地生物一樣都離不開微生物的作用。同樣地,海洋微生物對於淨化有機物污染,如石油、有機農藥等污染起積極的作用。水域的淨化離不開微生物。
越來越多的事實表明,海洋細菌不僅起著還原者的作用,而且還是許多種海洋動物的直接餌料。海洋細菌,無論是在水中和海底沉積物里,其生物量都相當可觀。據對大西洋一些淺水站位調查,細菌的生物量約占總的微小生物量的9.4%;而在大陸坡水中,細菌所占的比例增至52.5%。在某一個大洋站位,細菌的生物量在水中所占的比例高達94%。已查明,某些海洋浮遊動物的食物來源中細菌所占的比例,可達30~50%。
能流 海洋生物的能量轉換效率(生態效率)要比陸地生物的高,這是因為陸地植物所含的蛋白質比海洋浮游植物低得多。由浮游植物到植食性動物,生態效率約20%左右;由浮遊動物到浮遊動物捕食者,約15%;由低級肉食者到高級肉食者,約10%。
生物的進化,從單細胞到高等動物以至人類,都是沿著改善獲能效率和增加獲取能量的途徑進行的。在生物進化的過程中,生產者和消費者各自的進化水平是相匹配的。化石的分析結果證明,甲藻的出現時期與魚類的最盛時期相吻合,而硅藻的出現時間大體與鯨類的出現時間相同。J.H.賴瑟(1969)指出以硅藻為基礎的食物鏈要短些,而以鞭毛藻為基礎的食物鏈要長得多。鯨處於以硅藻為基礎的食物鏈上,因此能更好地取得能量,彌補其個體大因而能量消耗也大的不足。
雖然整個海洋是一個連續的整體,不同海域之間沒有明確的界線,但根據環境條件的差異,根據生物和環境的特點,可將海洋生態系統分為紅樹林、河口、淺海、潮間帶、上升流、大洋區生態系統,或浮游生物、底棲生物、游泳生物生態系統,或某某海區生態系統。目前有關生態系統的知識還很貧乏,研究工作比較集中於上升流、河口、紅樹林和幾個海區的生態系統。概括已有的資料,已建立幾個簡單的生態系統模型(圖6)。建立模型的目的,是為了更好地表達生態系統內部各種因素、成分之間的複雜關係,幫助人們預測在生態系統遭到各種壓力下可能發生的變化,從而達到管理生態系統的目的。
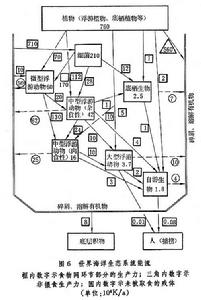 海洋生態
海洋生態深海生物能忍高壓。雖然有些淺海生物也能忍受較高的壓力,比如附著在潛水器表面的生物,如綠管滸苔、石蓴、總合草苔蟲、紫貽貝、布紋藤壺等,在潛水器下降到2000~3000米水深後仍然能存活。但根據生理學試驗,600個大氣壓對大多數淺海生物有致死作用。因此,從垂直分布來看,6000米深度似乎是個重要的分界線。曾有報告,中太平洋的深海溝中的125種動物,有77種是在6000米以上水層所沒有見到的。
與淺海生物比較,深海生物一般個體數量少,但種類數相對較多、多樣性高。對此有不同的解釋。H.L.桑德斯(1968、1973)認為,多樣性高是由於食物等競爭造成的。但有的學者卻認為捕食是關鍵。較多的調查結果表明,深海生物的多樣性僅僅發現在2000~3000米水深處,而5000~6000米以下的海底,生物的多樣性並不高。
為適應食物少和黑暗的環境,許多深海魚類,口部相對擴大,骨骼肌肉減少,且有發達的發光器官和結構。
深海生物一般代謝作用和生長都很慢。據估計,深海的貝類,長到8毫米大約需100年的時間。曾有一隻潛水器掉進深海中,經10個月後從1540米處打撈出來,發現放在桌上的三明治仍然完整無損,這表明細菌的作用非常緩慢,但也有例外。
1977年,美國伍茲霍爾海洋研究所曾用深海潛水器“阿爾文”號在加拉帕戈斯群島以東300公里,水深2500米處進行調查,調查區域是海洋板塊形成區,發現從地下噴出泉水,泉水口附近水溫高達20℃(沒有熱泉處的海水是2℃)。在噴出孔附近有豐富的生物群落。其中有個體30~40厘米的貝類,這種貝一年可長4厘米(約比其他深海底的貝類生長速度高500倍),將殼打開,可見到內有血紅蛋白(一般軟體動物是血藍蛋白)。還有一種具長棲管的須腕動物,管的直徑10厘米長可達30米。此外,還有許多腔腸動物、環節動物、甲殼動物。
深海底棲生物的食源可能包括由上層水中下沉的碎屑和溶解的有機物質,以及化能合成細菌通過氧化硫化氫取得能量而製造的有機物。目前認為,後者是最主要的來源。因為從地下噴出的熱水含有大量的硫化氫(30~40ppm),硫磺細菌利用氧化硫化氫所獲得的能量將水中CO2合成碳水化合物。海底硫磺細菌實際上起著與淺海水域光合植物相同的作用。即硫磺細菌是深海海底的生產者。這說明,生產者的能源不僅可來自太陽,而且還可來自地球的內部。這是一個重大的發現。根據這一論點,須腕動物的營養問題可得到解釋:這種動物沒有口,也沒有消化道,但在體內有大型的腔,稱為營養體,細菌在腔內大量繁殖,動物的觸手可吸收無機物供細菌之需,而細菌則合成有機物供動物之用。動物和細菌營互利共生關係。
