基本信息
由於須腕動物均深海生活,加上其特殊的結構,在20世紀之前還沒有被人發現,直到1900年才被法國動物學家M.Caullery在印度尼西亞深海區採到第一個須腕動物標本。從那以後,陸續被發現,1955年被蘇聯學者A.V.Ivanorv建立起一個獨立的門,到目前已報導了近百個種。其地理分布也逐漸擴大,從太平洋西北海岸到大西洋的西海岸均有發現,大多分布在大陸斜坡上150—1500m深處,最深的可達數千米。最近也發現少數種類生活在淺海(22m處),我國東海也有發現。

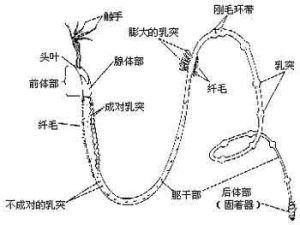 須腕動物
須腕動物其他信息
前體部之後為軀幹部,兩者之間有隔板。軀幹部上有各種不同形態的乳突(papillae),或單個排列,或成雙排列,或成堆排列。另外還有兩個剛毛狀環行纖毛帶(girdles),它們的作用是使蟲體在管內粘著,以及支持身體在管內上下運動。
軀幹部後為短小的後體部,是由5—23個具剛毛的體節組成,其結構及起源相似於環節動物。後體部可能用以附著身體,並通過管的下端開口用以在軟泥中挖掘。須腕動物很脆弱,以前採到的標本大都缺乏後體部,以致將須腕動物列入後口動物。直到1964年才採到完整的標本,知道了後體部的存在,這才確定了須腕動物與環節動物更相近,而被列入原口類動物。
須腕動物的體壁由角質層、單層上皮細胞、環肌及縱肌組成,每個體區內均有體腔囊,前體部的體腔伸入觸手中,後體部具分節的體腔囊。
須腕動物一個顯著的特徵是其完全缺乏消化系統。過去一直認為觸手的羽枝是其過濾及攝食的結構,並由羽枝分泌消化酶,進行體外及胞外消化。但近年來的研究沒有發現消化酶的存在,更多的傾向於須腕動物是靠直接吸收溶於水中的有機物顆粒而獲得營養。須腕動物沒有專門的呼吸系統,由體表、觸手及羽枝進行氣體交換。具閉管式循環系統,由背血管及腹血管組成。腹血管的前端膨大成心臟,由它給出血管到觸手及羽枝。身體後端有橫行血管相連。可能通過前體部的體腔管進行排泄,軀幹部的一些細胞也能貯存代謝物質。神經系統位於表皮內,前體頭葉背面神經細胞聚集形成腦,由腦發出神經到觸手及身體後端,也有巨大神經纖維。雌雄異體,在軀幹部兩側的體腔中有兩個長圓形生殖腺。雄性生殖孔位於軀幹的前端,可在輸精管的末端形成精莢。雌性的有兩個U形輸卵管,實際是體腔管,呈漏斗狀開口在軀幹的中段。關於精子的傳遞、受精及產卵過程尚未被觀察到,但在西伯達蟲中曾看到精莢通過觸手送到管口,可能是通過漂浮作用到達雌性個體。卵產於管中,並在其中發育。卵圓形,屬多黃卵,經螺鏇卵裂或放射卵裂、外包法形成原腸胚,直接發育成成蟲後離開母體。由管口收集的胚胎是蠕蟲狀,具卵黃,有纖毛帶。離開母體後沉入海底。
多數學者認為,須腕動物的分類地位與環節動物相近,但尚有爭議,有待進一步研究確定。
