對釀酒酵母代謝工程的利用
根據代謝工程原理,採取多拷貝整合策略,利用整合載體pYMIKP,將來自嗜熱細菌Thermusthermophilus的木糖異構酶(XI)基因xylA和釀酒酵母(Saccharomycescerevisiae)自身的木酮糖激酶(XK)基因XKS1,插入釀酒酵母工業菌株NAN27的染色體中,得到工程菌株NAN114。酶活測定結果顯示,NAN114中XI和XK的活性均高於出發菌株NAN27,表明外源蛋白在釀酒酵母工業菌株中得到活性表達。對木糖、葡萄糖共發酵搖瓶實驗結果表明,工程菌NAN114消耗木糖4.6g/L,產生乙醇6.9g/L,較出發菌株分別提高了43.8%和9.5%。首次在釀酒酵母工業菌株中建立了XI路徑的木糖代謝途徑。
共發酵研究
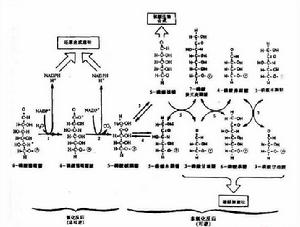 木酮糖激酶釀酒酵母(Saccharomycescerevisiae)是傳統的乙醇生產菌株,因缺乏將木糖轉化為木酮糖的代謝途徑而不能很好地利用木糖[1,3~5]。自然界中由木糖轉化為木酮糖的代謝途徑有兩條:其一,木糖經依賴NADPH的木糖還原酶(xylosereductase,XR)還原形成木糖醇,再由依賴NAD+的木糖醇脫氫酶(xylitoldehydrogenase,XDH)催化形成木酮糖[4]。其二,木糖在木糖異構酶(xyloseisomerase,XI)的作用下直接轉化木糖形成木酮糖[5]。同時,代謝途徑下游的木酮糖激酶(xylulokinase)催化木酮糖磷酸化形成5磷酸木酮糖的酶促反應也是木糖代謝的限速步驟之一[6,7]。由於XR和XDH催化反應需要的輔酶不同,因而在木糖代謝過程中造成氧化還原不平衡,直接導致副產物木糖醇的大量積累[4]。木糖異構酶可以直接轉化木糖到木酮糖,不需要輔酶,被認為是構建代謝木糖釀酒酵母工程菌株的便利途徑,但許多不同種屬來源的木糖異構酶在釀酒酵母中均沒有得到活性表達[5],直到1996年Walfridsson等[5]首次在釀酒酵母中實現了嗜熱細菌Thermusthermophilus木糖異構酶的活性表達。但是,Walfridsson等[5]採用的是附加體載體系統,以單倍體營養缺陷基因型的實驗室菌株作為受體菌,外源基因在重組菌株中的表達穩性較差,不具備產業化意義。對工業生產菌株的代謝工程改造是工程菌產業化的前提條件,釀酒酵母生產菌株多倍性及野生基因型的狀態,不能適用常規的分子生物學操作體系[8]。本文在建立了釀酒酵母工業菌株轉化體系的基礎上,以釀酒酵母酒精生產工業菌株NAN27為出發菌株,通過本實驗室構建的整合載體pYMIKP,將T.thermophilus的木糖異構酶基因xylA以及同源木酮糖激酶基因XKS1,整合於NAN27染色體中,並對工程菌株木糖葡萄糖共發酵進行了初步研究。
木酮糖激酶釀酒酵母(Saccharomycescerevisiae)是傳統的乙醇生產菌株,因缺乏將木糖轉化為木酮糖的代謝途徑而不能很好地利用木糖[1,3~5]。自然界中由木糖轉化為木酮糖的代謝途徑有兩條:其一,木糖經依賴NADPH的木糖還原酶(xylosereductase,XR)還原形成木糖醇,再由依賴NAD+的木糖醇脫氫酶(xylitoldehydrogenase,XDH)催化形成木酮糖[4]。其二,木糖在木糖異構酶(xyloseisomerase,XI)的作用下直接轉化木糖形成木酮糖[5]。同時,代謝途徑下游的木酮糖激酶(xylulokinase)催化木酮糖磷酸化形成5磷酸木酮糖的酶促反應也是木糖代謝的限速步驟之一[6,7]。由於XR和XDH催化反應需要的輔酶不同,因而在木糖代謝過程中造成氧化還原不平衡,直接導致副產物木糖醇的大量積累[4]。木糖異構酶可以直接轉化木糖到木酮糖,不需要輔酶,被認為是構建代謝木糖釀酒酵母工程菌株的便利途徑,但許多不同種屬來源的木糖異構酶在釀酒酵母中均沒有得到活性表達[5],直到1996年Walfridsson等[5]首次在釀酒酵母中實現了嗜熱細菌Thermusthermophilus木糖異構酶的活性表達。但是,Walfridsson等[5]採用的是附加體載體系統,以單倍體營養缺陷基因型的實驗室菌株作為受體菌,外源基因在重組菌株中的表達穩性較差,不具備產業化意義。對工業生產菌株的代謝工程改造是工程菌產業化的前提條件,釀酒酵母生產菌株多倍性及野生基因型的狀態,不能適用常規的分子生物學操作體系[8]。本文在建立了釀酒酵母工業菌株轉化體系的基礎上,以釀酒酵母酒精生產工業菌株NAN27為出發菌株,通過本實驗室構建的整合載體pYMIKP,將T.thermophilus的木糖異構酶基因xylA以及同源木酮糖激酶基因XKS1,整合於NAN27染色體中,並對工程菌株木糖葡萄糖共發酵進行了初步研究。相關詞條
拷貝、嗜熱細菌、酵母、乙醇、菌株、木糖醇脫氫酶
