
基本定義
 基因突變的未來新物種
基因突變的未來新物種對於植物而言,二倍體小麥與四倍體小麥之間雖然不能得到可育的後代,但它們屬於同一物種中的不同亞種。
物種研究
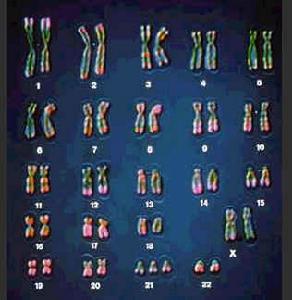 染色體
染色體研究生物分類的方法和原理的生物學分支。分類就是遵循分類學原理和方法,對生物的各種類群進行命名和等級劃分。分類學曾被稱為系統分類學,但它與系統學很易混淆,系統學是研究生物的分異度(多樣性)以及它們中間的任何一個類群和其他所有類群的各種關係的科學,曾稱為分類系統學。三者的共同目的是從理論上和實踐上,闡明種類之間的關係(或親緣關係),建立自然系統,確定各類群的命名和排序,總結其進化歷史。
地球上現生的物種以百萬計,千變萬化,各不相同,如果不予分類,不立系統,便無從認識,難以利用。分類系統是生物種類的查找系統,可藉以認識和查取有關資料。分類的對象是形形色色的種類,都是進化的產物;分類學在於闡明種類之間的歷史淵源,使建立的分類系統反映進化歷史。因而從理論意義上說,分類學是生物進化的歷史總結。
分類學是綜合性學科。生物學的各個分支,從古老的形態學到現代分子生物學的新成就,都可吸取為分類依據。分類學亦有其自己的分支學科,如以染色體為依據的細胞分類學(或染色體分類學),以血清反應為依據的血清分類學,以化學成分為依據的化學分類學,等等。動物、植物和細菌,作為三門分類學,各有其特點;病毒分類則尚未正式採用雙名制和階元系統 。
分類學歷史
 物種起源
物種起源古希臘哲學家亞里士多德採取性狀對比的方法區分物類,如把熱血動物歸為一類,以與冷血動物相區別。他把動物按構造的完善程度依次排列,給人以自然階梯的概念。
17世紀末葉,英國植物學者J.雷曾把當時所知的植物種類,作了屬和種的描述,所著《植物研究的新方法》(1682)是C.林奈以前的一本最全面的植物分類總結。J.雷還提出“雜交不育”作為區分物種的標準。
近代分類學誕生於18世紀,它的奠基人是瑞典植物學者林奈。林奈為分類學解決了兩個關鍵問題:①建立了雙名制,每一物種都給以一個學名,由兩個拉丁化名詞所組成,第一個代表屬名,第二個代表種名。例如Pantheratigris是虎的學名,Bombyxmori是蠶的學名。②確立了階元系統。林奈把自然界分為植物、動物和礦物三界,在動植物界下,又設有綱、目、屬、種四個級別,從而確立了分類的階元系統。每一物種都隸屬於一定的分類系統,占有一定的分類地位,可以按階元查對檢索。並首先在1753年印行的《植物種志》和1758年第10版《自然系統》中套用於植物和動物。這兩部經典著作,標誌著近代分類學的誕生。林奈相信物種不變,他的《自然系統》沒有親緣概念,其中6個動物綱是按哺乳類、鳥類、兩棲類、魚類、昆蟲、蠕蟲的順序排列的。J.-B.de拉馬克把這個顛倒了的系統撥正過來,從低級到高級列成進化系統。他還把動物區分為脊椎動物和無脊椎動物兩類,並沿用至今。但是,由於他的進化觀點在當時沒有得到公認,因而對分類學影響不大。直到1859年,C.R.達爾文的《物種起源》出版以後,進化思想才在分類學中得到貫徹,明確了分類研究在於探索生物之間的親緣關係,使分類系統成為生物系譜——系統分類學由此誕生。
 林奈
林奈種和屬的學名後常附命名人姓氏,以標明來源,便於查找文獻。表上家蠶,蠶蛾屬學名後的L.,是命名人林奈Linnaeus的縮寫,馬尾松學名後的Lamb.,是命名人蘭伯特Lambert的縮寫,通常除林奈等少數之外,一般命名者姓名引用時不應縮寫。亞種命名採取3名制,即在屬名、種名後再加亞種名。例如華南虎和東北虎都是虎的亞種,前者的學名是PantheratigrisamoyensisHilz-heimer,後者是PantheratigrisaltaicaTemminck。變種學名亦來取3名制,只是在種名與變種之間加上拉丁文變種varietas的縮寫var.,例如中華按蚊原先鑑定為赫坎按蚊的變種(現已分出為獨立的種)學名曾是Anopheleshyrcanesvar.sinensisWedemann。
分類名稱要求穩定,一個屬或種(包括種下單元)只能有一個學名,如果有兩個或多個時,便是“同物異名”,必須於其中核定最早提出的學名,摒棄較後的異名。一個學名只能用於一個對象(屬或種),如果有兩個或多個對象者,便是“異物同名”,必須於其中核定最早的命名對象,而其他的同名對象則另取新名。這叫做“優先律”,動物和植物分類學界各自製訂了《命名法規》,所以在動物界和植物界間不存在異物同名問題,如Cystiphyllum既是褐藻又是古生代珊瑚的屬名。“優先律”是穩定學名的重要措施。優先律的起始日期,動物是1758年,植物是1820年,細菌則起始於1980年1月1日。
法規的另一重要規定是模該方法。凡發表新種(包括新屬和新亞種),必須指定一個標本作為命名的模式標本,叫做“正模”,簡稱模本。“副模”是原描述所根據的其他標本,特別是和正模同時同地採得的標本;“配模”是副模中選取的與正模異性的標本。在模本遺失或未定的情況下,可以選取合適標本作為“新模”,以代替正模,一般要從原產地和原層位採集。與模本產地相同而在以後採得的標本可立為“地模”。模式標本是鑑定的對照標本。
鑑定學名是取得物種有關資料的手段,即使是前所未知的新種類,只要鑑定出其分類隸屬,亦可預見其一定特徵。分類系統是檢索系統,也是信息存取系統。許多分類著作,如基於區系調查的動植物志,記述某一國家或地區的動植物種類情況,作為基本資料,都是為鑑定、查考服務的 。
概念與標準
 物種
物種在林奈時代,人們相信物種是不變的,同種個體符合於同一“模式”。模式概念(不同於模式標本),淵源於古希臘哲學的古老的概念,套用到整個分類系統,概念假定所有階元系統中的各級物類單元,都各自符合於一個模式。對神創論者來講,由各級模式所組成的物類階元,正好說明了“造物主”的智慧設計。模式概念亦是形態學概念,所謂模式是指形態模式——物種是由形態相似的個體所組成的,同種個體符合於同一形態模式。這是物種的形態學定義,也是形態學標準。另一物種標準是雜交不育。早在林奈以前,J.雷就明確指出,物種是繁殖單元,同種個體可以交配繁殖,異種之間則雜交不育,形成生殖隔離。生殖隔離和模式概念一樣,符合於物種不變的觀點,因為它突出了種間間斷,無視歷史連續。林奈時代的物種概念,包含兩個基本內容:①不變,②客觀存在。由於不變,而且是“上帝”所創造,物種當然是客觀存在。種內形態同一,種間生殖隔離則是物種不變的兩個標準。
達爾文的物種概念與此相反,它的基本內容是:①變,②人為單元。進化論說明了物種是變的,變化中的物種否定了自身的存在。達爾文在《物種起源》內說:“可見‘物種’這名詞,我認為是為了方便起見,任意地用來表示一群很相似的個體的,它在本質上和‘變種’沒有區別”。達爾文不考慮物種定義,因為物種既是人為單元,便不可能有客觀標準,更不需要定義。他的進化論證明了種間的歷史連續,卻忽視了種間的間斷意義。
物種之變與不變曾經是進化論和特創論的鬥爭焦點,是勢不兩立的觀點。但是,分類學的事實說明,每一物種各有自己的特徵,沒有兩個物種完全相同;而每個物種又保持一系列祖傳的特徵,據之可以決定其界、門、綱、目、科、屬的分類地位,並反映其進化歷史。整個階元系統內各級物類之間的特徵關係,表現為祖征與新征的對立統一。界的特徵,如細胞壁、葉綠體和光合作用之作為植物特徵,對界下單元(門、綱、目、科、屬、種)來講,是祖征;而在界間對比,如植物界與動物界或真菌界對比,則又表現為各界的新征。界下的各級單元,從門到種,亦都如此。
 達爾文
達爾文分類工作的基本內容是區分物種和歸合物種,前者是種級和種下分類,後者是種上分類。種群概念提高了種級分類水平,改進了種下分類,其要點是以亞種代替變種。亞種一般是指地理亞種,是種群的地理分化,具有一定的區別特徵和分布範圍。亞種分類反映物種分化,突出了物種的空間概念。含有兩個或多個亞種的物種稱多型種,不分亞種的物種稱單型種。形態十分相似但生殖上隔離的物種稱姐妹種,又稱隱種。分布區域重疊的物種稱同地種,分布不重疊的物種稱異地種。地理亞種都是異地亞種,在交接區域常常有中間類型。
變種這一術語過去用得很雜,有的指個體變異,有的指群體類型,意義很不明確,在動物分類中已廢除不用。在植物分類中,一般用以區分居群內部的不連續變體。生態型是生活在一定生境而具有一定生態特徵的種內類型,常用於植物分類。人工選育的動植物種下單元稱為品種。
由於種內種間變異錯綜複雜,分類學者對種的劃分有時分歧很大。根據外部形態的異同程度作為劃分物種依據而劃分的稱為形態種,由於對各種形態特徵的重要性認識不一,使劃分的種因人而異,尤其是分類學者對某些特徵的“加權”常使它們比其他特徵更具重要性,而造成主觀偏見。古生物種或化石種中的大多數是以形態特徵為依據,甚至以硬組織,如外殼、骨骼等部分形態特徵為依據而建立的,應歸於形態種的範疇。植物的各個部分常單獨保存為化石,某些不屬於同一科的生物,由於單獨保存的葉、根等,很難確立其系統關係,僅以形態相似而建立的屬為形態屬,如櫛羊齒(Pecopteris)等;其單獨保存的器官可構成器官屬,如鱗木類的孢子葉化石叫鱗孢葉屬(Lepidostrobophyllum),孢子葉穗化石稱鱗孢穗屬(Lepidostrobus),而根座常被稱為Stigmaria。
另外,強調染色體數目(如二倍體、多倍體、無融合生殖種等)作為分種依據而將外部形態作為次要特徵依據的稱為生物學種。植物分類有大種和小種之分:“歸併派”主張大種概念,又稱“林奈種”,把種的範圍劃得較大,有時甚至很大,包括近緣種在內;“細分派”主張小種概念,又稱“約爾丹種”,常把亞種、變種、生態型等劃分為種,種下不再分類。動物分類學亦有歸併和細分兩派,但不作大種和小種的區分。歸併派和細分派的存在,說明了分類學家對物種標準還缺少一致的認識。作為動物種,不同種間不能繁殖,但作為植物種則可以雜交而產生有繁殖能力的後代,如小麥。因此傾向於把二倍體作為種的代表。
這一切說明物種的概念及定義並未真正解決,同時,種上分類亦同樣有歸併與細分之爭,例如細分派把貓科分為28個屬,而極端的歸併派只承認一個屬。
系統原理和分類方法
一個物種或物類,以至整個植物界和動物界,都有自己的歷史。研究系統發育就是探索種類之間歷史淵源,以闡明親緣關係,為分類提供理論依據。儘管在分類學派中有綜合(進化)分類學、分支系統學和數值分類學3大流派,但在其基本原理上都有許多共同之處,不過各自強調不同的方面而已。系統分類的要求是:①共同起源。每一個物類都有一個起源的問題,要求同一系統內的物種或物類必須是起源於共同的祖先的,這樣才能反映自然系譜。這叫做建立單系系統,單系是指一個物類起源於一個祖種。復系則指起源於兩個或多個祖種。分類學要求建立單系系統,不取復系系統。③分支發展。一個新物類的產生最初總是少數,在理論上該是一個物種。從這個最初的祖種,通過分支發展,從少到多,形成為許多不同支系。系統發展是分支發展的過程,分支關係反映親緣關係。圖1是簡化的系統樹,從A到H代表8個物種或物類,A和B的共同祖先在Ⅰ處,A和C的在Ⅱ處,A和F的在Ⅲ處。根據這種分支關係,W.亨尼希對親緣關係下了一個定義:當物種A和B來源於某一個共同祖先,而這個祖先卻不是物種C的祖先時,則A和B的關係要比A和C為近。因此,共同祖先的遠近,決定親緣關係的遠近。他指出,所謂單系類群“是一些或大或小的類群,其群內的物種之間的相互關係比群外的任何物種為近”。例如,圖1中AB、CD、FG、CDE、FGH、ABCDE都是這樣的單系類群,而BCDE由於A的存在,DE由於C的存在,GH由於F的存在,都不是單系類群。這是亨尼希的、亦即分支系統學的嚴格的單系概念。③級進發展(或級序發展)。分支是橫的分化,級進是縱的上升,是生物從低級到高級的發展。分支與級進是相輔相成的。例如:硬骨魚類在分支發展中有一支演進為兩棲類,兩棲類在分支中有一支演進為爬行類,另一支演進為哺乳類,而鳥類和哺乳類,又再各自分支發展。所以分支中有級進,級進後再分支,系統發育是分支與級進的統一發展過程(分支與級進是相對的,大分支中有大級進,小分支中有小級進)。
假定圖1上的BCDE代表爬行動物的4個支系,A是由此分出的鳥類分支,按照階段級進的觀點,鳥類和爬行類的四支比較,代表進化的一個新的階段,必須分列為獨立的一個生物群,以與其他4支相區別。這就是一般所採取的鳥綱與爬行綱並立的傳統分類,也就是所謂進化分類學的分類。進化分類學派認為這個分類並不違反單系系統的原則,因為鳥類(A)作為一個單元,源出於同一祖種,而爬行類的四個支(BCDE)作為另一單元,同樣是出於一個祖種。分支學派把系統圖上從同一基點所分出的支系稱為姊妹群,圖1上如Ⅰ處分出的A和B兩支,Ⅱ處分出的AB和CDE兩支,以及Ⅲ處分出的ABCDE和FGH兩支都是姊妹群。姊妹群在分類上應列為同等級別,如果圖上A代表鳥類,BCDE代表爬行類4個支系,則A支應與B支同級,不能與BCDE四支同級。
這是當前系統分類學上的兩個學派的分歧:分支系統學派以宗譜分支為衡量親緣的唯一標準;進化分類學派則考慮兩個標準,宗譜分支和進化水平(階段改進)。分支與級進交錯發展,因而在分類工作中經常會遇到一個和兩個標準的分歧,成為兩個學派的爭論交點。
特徵對比是分類的基該方法。所謂對比是異同的對比:“異”(差異性)是區分種類的根據,“同”(相似性)是合併種類的根據。分析分類特徵,首先要考慮反映共同起源的共同特徵。但有同源和非同源的不同。例如鳥類的翼和獸類的前肢是同源器管,可以追溯到共同的祖先,是“同源特徵”。恆溫在鳥獸是各別起源,並非來自共同祖先,是“非同源特徵”。系統分類採用同源特徵,不取非同源性狀。亨尼希把同源特徵分為祖征和裔征兩類:祖征是來自較早祖先而在後代保持不變的特徵;裔征是兩個姊妹支系來自最近祖先的派生特徵,不見於較早祖先。每一物種或物類都是祖征和裔征鑲嵌,共同祖征是追溯歷史淵源的根據,共同裔征是建立單系的根據。進化分類學派亦要求採用同源特徵,但不作特徵分類。他們從共同性探索共同祖先,又從差異性衡量進化水平。上面提到鳥類和爬行類的問題,已知鳥類和鱷類最近緣,按照分支學派的分析,兩者具有共同的裔征(如內鼻孔具次生齶,其他爬行類內鼻孔無次生齶),在系譜上是一對姊妹支系,組成為一個單系群。但進化分類學派重視特徵差距,他們根據鳥類具有羽毛、前肢特化為翼以及恆溫等特徵,認為兩者差距很大,反映不同的進化階段,主張鳥類應與鱷目分開,而與爬行類(包括鱷目)並立為綱。這是兩個學派對特徵衡量的分歧,也就是宗譜分支與進化水平的分歧。
分類特徵反映進化歷史,系統分類要求從歷史觀點衡量特徵價值。但特徵衡量常會出現分歧,重視這一或那一特徵,會得出不同的分類。在某些情況下,由於同源和非同源不易區分,衡量又會陷入循環論,如:共同的特徵反映共同祖先,共同的祖先產生共同特徵。基於這些情況,為了促進分類的穩定性和重複性,數量分類學主張“總體相似性”的觀點,其方法是:採用大量能以數值表示的特徵,編成符號,運用電子計算機。求得相似性係數,作出分類。數量分類不先考慮親緣,但得出的結果在很多情況下和傳統分類近似。
生命現象的各個方面都可為分類提供特徵,最常用的是形態,尤其是外部形態。但目前趨向愈來愈重視生理、生化、遺傳等方面的特徵,如DNA(脫氧核糖核酸)的含量比較,蛋白質的成分分析,染色體的祖型分析,以及植物化學成分、動物交配行為等等。如以蟋蟀鳴聲為特徵,使原先單憑形態特徵所未能區分的近緣種類得以鑑別。
生物的界級分類
林奈把生物分為兩大類群:固著的植物和行動的動物。兩百多年來,隨著科學的發展,人們逐漸發現,這個兩界系統存在著不少問題,但直到20世紀50年代,仍為一般教本所遵從,基本沒有變動。最初的問題產生於中間類型,如眼蟲Euglena綜合了動植物兩界的雙重特徵,既有葉綠體而營光合作用,又能行動而攝取食物。植物學者把它們列為藻類,稱為裸藻;動物學者把它們列為原生動物,稱為眼蟲。中間類型是進化的證據,卻成為分類的難題。為了解決這個難題,早在19世紀60年代,人們建議成立一個由低等生物所組成的第三界,取名為原生生物界(Protista),包括細菌、藻類、真菌和原生動物。這個三界系統解決了動植物界限難分的問題,但未被接受,整整100年後,直到20世紀60年代,才開始流行了一段時間,為不少教科書所採用。
生命的歷史經歷了幾個重要階段,最初的生命應是非細胞形態的生命,當然,在細胞出現之前,必須有個“非細胞”或“前細胞”的階段。病毒就是一類非細胞生物,只是關於它們的來歷,是原始類型,還是次生類型,仍未定論。從非細胞到細胞是生物發展的第二個重要階段。早期的細胞是原核細胞,早期的生物稱為原核生物(細菌、藍藻)。原核細胞構造簡單,沒有核膜,沒有複雜的細胞器。從原核到真核是生物發展的第三個重要階段。真核細胞具有核膜,整個細胞分化為細胞核和細胞質兩個部分:細胞核內具有複雜的染色體裝置,成為遺傳中心;細胞質內具有複雜的細胞器結構,成為代謝中心。由核質分化的真核細胞,其機體水平遠遠高出於原核細胞。從單細胞真核生物到多細胞生物是生命史上的第四個重要階段。隨著多細胞體形的出現,發展了複雜的組織結構和器官系統,最後產生了高級的被子植物和哺乳動物。
植物、菌類和動物組成為生態系統的3個環節。綠色植物(真核植物和原核藍藻)是自養生物,是自然界的生產者。它們通過葉綠素進行光合作用,把無機物質合成有機養料,供應自己,又供應異養生物。菌類(細菌和真菌)是異養生物(自養細菌除外),是自然界的分解者。它們從植物得到食料,又把有機食料分解為無機物質,反過來為植物供應生產原料。動物亦是異養生物,它們是消費者,是地球上最後出現的一類生物。即使沒有動物,植物和菌類仍然可以存在,因為它們已經具備了自然界物質循環的兩個基本環節,完成循環過程中合成與分解的統一。但是,沒有動物,生物界不可能這樣豐富多彩,更不可能產生人類。植物、菌類和動物代表生物進化的3條路線或3大方向。
以上是目前對生物進化的階段和方向的認識,是現代界級分類的根據。
當前最流行的分類是一種五界系統。五界系統反映了生物進化的3個階段和多細胞階段的3個分支,是有縱有橫的分類。它沒有包括非細胞形態的病毒在內,也許是因為病毒系統地位不明之故。它的原生生物界內容龐雜,包括全部原生動物和紅藻、褐藻、綠藻以外的其他真核藻類,包括了不同的動物和植物。不少學者認為,不必成立原生生物界,而把藻類和原生動物分別劃歸植物界和動物界,成為比較緊湊的四界系統。反之,亦還有不少人主張擴大原生生物界,把真菌劃歸在內,成為另一種四界系統。本卷採用六界系統。因為任何生物系統都應把病毒(連同類病毒)包括在內。病毒是非細胞生物,但不一定代表生命的前細胞階段。六界分類系統,在病毒界和細菌界之間還加有問號;因為病毒如果是原始生物,細菌作為早期的原核生物,很可能起源於“原病毒”。原核生物的最早化石記錄,包括細菌和藍藻,見於30多億年前的地層中;真核生物的最早記錄,包括金藻和綠藻,見於14~15億年前的地層中。
自養的生產者藍藻和異養的分解者細菌組成為原核時代的兩大生態環節,正如植物、真菌和動物組成為真核生物的三大生態環節。這五類生物,基於它們的重要時空地位,都應獨立為界,連同病毒,綜合為六界分類系統。
藍藻顯然起源於細菌,是細菌的一支;真核植物大概起源於原核藍藻,是藍藻的一支(見細胞起源)。從真核植物又分支發展為真菌和動物。一般說來,植物細胞有細胞壁和葉綠體,真菌細胞有壁無葉綠體,動物細胞無壁無葉綠體。如果把原核的藍藻和動植物的中間類型裸藻都歸在植物界內,列為藍藻門和裸藻門。真菌和動物的中間類型粘菌,以及真菌和藻類共生的地衣,歸在真菌界內,則可成為一種四界系統。
地方物種保護
改革開放是國家發展經濟的重大戰略決策。發展經濟不能忽略本土物種的保護性。本民族和本土物種的純正關係到自然界物種的生物鏈延續。隨著人類社會的科學化發展,以及鄉村城市化建設和人類轉基因工程引進範圍的不斷擴大,地方原始性物種的保護已迫在眉睫。近年來,由於中國城市化的建設規模和鄉村城市化的速度日新月異,引進非本土植物種類和基因改良的物種大量增長,原始地方物種數量不斷削減或絕種。同時,轉基因工程技術不斷深入,保護部分物種的純正繁衍成為人類社會發展的最為關鍵的研究課題。由於人類生產活動加速導致的原始物種的毀滅比自然淘汰率高1000倍之多。這個速度也超過了新生物種的進化。一、快速削減的地方區域性主要物種可分為幾大類:
⑴林地或非林地區域的昆蟲、爬行動物以及鳥類。
⑵海洋及地方水生生物類。
⑶原始野生植物類。
⑷微生物類。
就目前來看,隨著人類活動區域的擴大,原始林地和原始地貌遭到了嚴重的破壞,各國政府已放棄了對物種保護的承諾。生物可分為動物和植物兩大種類。地球上的動物和植物以及微生物的種類至少有600萬至1400萬種,也有一些人認為在3000萬種左右。被確認的物種大約在175萬種左右。大量的稀有物種分布在濕地和熱帶雨林以及海洋中。
 物種
物種⑴林地的大量砍伐。
⑵水質的化學污染。
⑶空氣中有毒有害氣體的污染。
⑷區域性大範圍物種的引進和自然雜交,形成非純正性物種的轉基因。
⑸人類轉基因工程造成的區域性純正物種的改變。
⑹工業化城市以及現代化城市範圍的擴大。
⑺原始林地以及原始地貌資源的大量開採。
⑻人類戰爭引發的毀滅性物種滅絕,比如,核子彈、化學武器的使用等。
⑼海洋水生生物的大量捕撈。
⑽能源開採過程中的泄漏,如,海洋石油的泄漏污染。
⑾化工原料的泄漏以及化學原料對土地和空氣的污染。
⑿殺蟲劑、滅草劑等農業化學藥品的大量使用。
⒀工業的快速發展帶來的綜合性有毒有害物質的形成。
⒁人為的物種滅殺。
⒂大氣變暖導致的海平面上升形成的陸地被淹。
⒃自然災害對物種的毀滅。
⒄大氣臭氧層空洞造成的紫外線輻射。
⒅氣候變暖造成的地球兩極冰川融化。
⒆氣候變暖產生的大氣環流改變,形成區域性的重大災害。
三、地方區域性物種的保護措施
 物種
物種⑴地方政府建立健全地方物種保護機構。
⑵制定地方物種保護法。
⑶加大地方物種保有量的宣傳力度,讓民眾知道地方物種的分類。
⑷設立地方物種保護園區,建立地方物種博物館。
⑸物種保護從娃娃學起,編制地方物種種類普及教材。
⑹建立物種保護監督機制。
⑺縮減城市建設規模。
⑻加大對化工以及有害物體排放企業的監督管理。
⑼縮減汽車製造業生產規模,加大汽車尾氣排放的檢測範圍。
⑽加大低碳工業的發展規模。
⑾嚴格實施生產企業的法律性保障措施。
⑿縮減對自然資源的過度性開發。
⒀禁止生活垃圾的焚燒和填埋處理,科學建立垃圾分類綜合性消納處理場地。此項工程是“利在當代,功在千秋”的民生工程。
四、不能以物種滅絕為代價謀求經濟成長點
對環境的保護不是一兩代人的事情,而是一個民族、一個國家的長期宏偉目標。由於地方政府追求區域性GDP因素和近年來的城市化發展速度過塊,對原始區域中的林地、地貌、水域、礦產等資源進行破壞性掠奪,導致地方原始生態的低谷性演變,大部分地方物種瀕臨滅絕。雖然經濟成長了,自然生態鏈卻遭到了嚴重的破壞,如果繼續發展下去,人類的自然生存空間將成為人類的墳墓,多發性和重大的自然災害將接踵而來。各國的政要應當遵守諾言,保護大自然,保護人類賴以發展的自然狀態。僅依靠幾個科學家、學者的呼聲是不夠的。要解決這個問題就要統一思想,整合全人類的文化要素精髓,這是毋庸置疑的。
