當DNA進行複製時,雙螺鏇結構解開成兩條單鏈,各自作為模板合成與之互補的新鏈。在子代DNA雙鏈中,一條是來自於親代,另一條完全重新合成。
DNA的複製
真核細胞的DNA複製有6大特點
1.複製的方式的半保留複製。
半保留複製:以DNA分子的雙鏈為模板,以鹼基互補配對為原則,合成一條新鏈的過程。新和成的DNA分子鏈中一條來自親本,另一條是新合成的。
2.複製是以一個起始點開是的,一個DNA分子能有一個或者多個這樣的起始點。即從一個點開始解鏇,複製。
3.複製是單向或雙向的,原核生物複製一般為單向,真核生物複製一般為雙向。
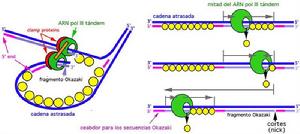 岡崎片段
岡崎片段4.複製起始時,需要一小段RNA引物激活DNA聚合酶的活性,促使多核苷酸鏈不斷增長。
5.複製的方向是有5端到3端添加核苷酸。
6.DNA的複製在一條鏈上是相對連續的,而另一條鏈是間斷的,先合成片段,再連成整體,即是DNA的合成2條鏈的複製不是同步的。合成的片段叫做岡崎片段。
半保留複製
半保留複製(semiconservative replication):一種雙鏈脫氧核糖核酸(DNA)的複製模型,其中親代雙鏈分離後,每條單鏈均作為新鏈合成的模板。因此,複製完成時將有兩個子代DNA分子,每個分子的核苷酸序列均與親代分子相同,這是1953年沃森(J.D.Watson)和克里克(F.H.C.Crick)在DNA雙螺鏇結構基礎上提出的假說,1958年得到實驗證實。
DNA既然是主要的遺傳物質,它必須具備自我複製的能力。瓦特森和克里克(1953)在提出DNA雙螺鏇結構模型的同時,對DNA複製也進行了假設。他們根據DNA分子雙螺鏇結構模型,認為DNA分子的複製,首先是從它的一端氫鍵逐漸斷開。當雙螺鏇的一端已拆開為兩條單鏈時,各自可以作為模板,從細胞核內吸取與自己鹼基互補的游離核苷酸(A吸取T,C吸取G),進行氫鍵的結合,在複雜的酶系統的作用下,逐漸連線起來,各自形成一條新的互補鏈,與原來模板單鏈互相盤鏇在一起,兩條分開的單鏈恢復為雙鏈DNA分子,與原來的完全一樣。DNA的這種複製方式稱為半保留複製(semiconservative replication),因為通過複製所形成的新的DNA分子,保留原來親本DNA雙鏈分子的一條單鏈。
DNA在活體內的半保留複製特徵已為1958年以來的大量試驗所證實。DNA的這種複製方式對保持生物遺傳的穩定具有非常重要的作用。
還可能存在其他兩種複製方式,都以原來親本DNA雙鏈分子作為模板鏈。一種方法稱為全保留複製(conservative replication),在複製過程中新的DNA分子單鏈結合在一起,形成一條新的DNA雙鏈,而親本DNA雙鏈仍然被保留在一起。另一種方法稱為散布式複製(dispersive replication),在複製過程中親本DNA雙鏈被切割成小片段,分散在新合成的兩條DNA雙鏈分子中。
1953年J.D.Watson和 F.H.C. Crick在提出DNA雙螺鏇結構時,對其互補關係予以很大的重視,而且提出了DNA的複製模型。DNA在進行複製時各以雙鏈中的每一條鏈作為模板,各個和互補的前體單核苷酸配對重合而形成與這二條單鏈各各對應的雙重子螺鏇二條。所謂互補就是指腺嘌呤一定只與胸腺嘧啶配對,鳥嘌呤一定只與胞嘧啶配對,新的單核苷酸排列在模板上時,其排列法是依原來鏈上的鹼基通過互補來決定的。這樣無論子分子與子分子間,還是子分子與母分子間,鹼基排列順序是完全相同。這樣一來具有和親本完全一樣的遺傳信息的子分子自我增殖了二倍。這時所產生的子雙重螺鏇分子一條鏈是從親代原封不動的接受下來的,只有相對的一條鏈是新合成的,所以把這種複製方式稱作半保留複製。這個模型曾用重同位素標記的DNA以密度梯度離心法進行分析,或用放射性同位素標記的DNA以放射自顯影法進行測定等等,用幾種不同原理的方法,曾在從人到病毒的許多種生物中進行了驗證,肯定了這個模型的正確性和普遍性。關於DNA是以半保留方式複製這一點已被認為是生物學中最基本的肯定性原理。
