
簡介
DNA複製過程中,2條新生鏈都只能從5端向3端延伸,前導鏈連續合成,滯後鏈分段合成,這些分段合成的新生DNA片段稱岡崎片段,細菌岡崎片段長度1000-2000核苷酸,真核生物岡崎片段長度100-200核苷酸.在連續合成的前導鏈中,U-糖苷酶和AP內切酶也會在錯配鹼基U處切斷前導鏈,任何一種DNA聚合酶合成方向都是從5'向3'方向延伸,而DNA模板鏈是反向平行的雙鏈,這樣在一條鏈上,DNA合成方向和複製移動方向相同(前導鏈),而在另一條模板上卻是相反的(後滯鏈)。那么在複製叉中新鏈是如何合成的呢?1968年岡崎(Okazaki)及其同事進行了一系列實驗,回答了這一問題。試驗檢測
 岡崎片段
岡崎片段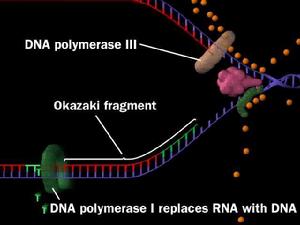 岡崎片段
岡崎片段由於這個實驗的結果使得人們普遍認為:無論是前導鏈還是後滯鏈都是先合成小片段,然後在連成大片段,稱為“不連續複製“(discontinuousreplication)。然而由模型預測應該只有一半放射標記存在於小片段中,但實驗的結果卻全部是小片段DNA,為什麼從前導鏈模板的3′-OH端延伸會不連續合成呢?人們在思考這個問題。1978年Olivera提出了半不連續(semidiscontinuous)複製模型,也就是說前導鏈上的合成是連續的,只有後滯鏈上的合成才是半連續的。現在已經弄清原來是由於細胞內都存在有dTTP和dUTP,而DNApolⅢ卻並不能區分它們,因此也會將dUTP加入到DNA中,形成A•U對。那么在DNA中為什麼沒有U的存在呢?這是因為E.coli細胞里有雙重“保險”,防止了U的“混入”。
第一道關是細胞里的dUTPase,它能使dUTP變成dUMP,dUMP是不能作為DNA合成的底物,這樣它就不再能加入DNA中。但總還有些漏網之“魚”,它們還是逃過此關,混入DNA中,這就靠第二道關來清除“異己”,這道關的主角是尿嘧啶N-糖苷酶(uracilN-glycosylase),它可以切斷混合尿苷的糖苷鍵,形成無Pu和Py位點(apurinicorapyrimidinic,AP),再由AP內切酶在AP位點切出一個缺口,進一步進行切除修復。在細胞內尿嘧啶N-糖苷酶作用較快,而AP酶作用較慢,在新鏈合成之初約1200bp就有可能摻進一個U,但很快就被尿嘧啶N-糖苷酶切斷糖苷鍵,在AP酶未作用前在脈衝標記實驗中就提取了,用NaOH沉澱時,AP位點十分易斷裂,所以前導鏈也成了小片段。以下實驗證實了這個解釋:
(1)在dut-突變體(dUTPase缺失)中岡崎片段比在dut+中為短。這是因為U摻入機會增加;(2)在ung-(尿嘧啶N-糖苷酶缺失)突變體中,新合成的DNA約有一半由片段組成。(3)因為尿嘧啶N-糖苷酶缺失,不會切除U的糖苷鏈,也就不會出現AP位點,所以鹼沉澱時不易斷裂,從而保持了半不連續的原貌。
在dut-,ung-雙突變體中,結果和實驗(2)相同,更進一步證實了此推測。真核岡崎片段的成熟機制比較複雜,由Polδ、RNaseHI、FEN1、Dna2和DNA連線酶等多種蛋白質協同通過兩種機制加工完成。Polδ的3′→5′核酸外切酶活性能替代FEN1,在岡崎片段成熟中通過阻止過量的DNA鏈替代合成,保證產生能被DNA連線酶連線的具有單一切口兩個DNA片段,有利於保持基因組的穩定性。
DNA雙鏈
在DNA雙鏈進行半保留複製時,在複製點附近新合成的與親代DNA鏈互補的DNA片段。是岡崎令治等(1966)首先發現的。在大腸桿菌約為1千-2千核苷酸的長度。DNA聚合酶只使脫氧核苷酸在5′→3′的方向重合。而半保留複製在5′→3′方向重合的同時,在3′→5′方向的重合也是不可缺少的。此矛盾由岡崎片段的發現,以及在其基礎上的不連續複製模式而解釋了。在複製點,如圖所示,首先由DNA聚合酶的作用,在5′→3′方向與親代DNA分子雙方的鏈互補,由脫氧核苷酸配對重合,形成短的DNA片段,它們再由DNA連線酶的作用而結合起來形成長的DNA分子,並逐漸複製成為DNA雙鏈。
遺傳信息
 岡崎片段
岡崎片段(一)DNA的半保留複製
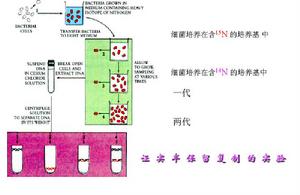
Waston和Click在提出DNA雙螺鏇結構模型時曾就DNA複製過程進行過研究,他們推測,DNA在複製過程中鹼基間的氫鍵首先斷裂,雙螺鏇解鏇分開,每條鏈分別作模板合成新鏈,每個子代DNA的一條鏈來自親代,另一條則是新合成的,故稱之為半保留式複製(semiconservativereplication)。
1958年Meselson和Stahl進行了如圖8-3-5的實驗證明了DNA分子是以半保留方式進行自我複製的。
(二)DNA複製的起始,方向和速度
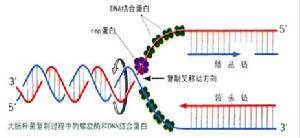
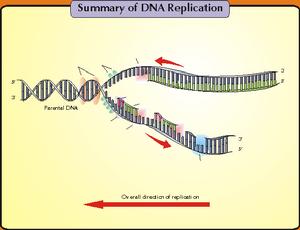
DNA在複製時,雙鏈DNA解鏇成兩股分別進行。其複製過程的複製起點呈現叉子的形式,故稱複製叉。以複製叉向前移動的方向為標準,一條模板鏈為3’—〉5’走向,在其上DNA能以5’—〉3’方向連續合成,稱為前導鏈(leadingstrand);另一條模板鏈為5’—〉3’走向,在其上DNA也是5’—〉3’方向合成,但與複製叉移動的方向正好相反,故隨著複製叉的移動形成許多不連續的岡崎片段,最後在連成一條完整的DNA鏈,該鏈稱為後隨鏈(laggingstrand)。實驗證明DNA的複製是由一個固定的起始點開始的。一般把生物體的單個複製單位稱為複製子。一個複製子只含一個複製起點。一般說,細菌,病毒即線粒體DNA分子均作為單個複製子完成其複製,真核生物基因組可以同時在多個複製起點上進行雙向複製,即它們的基因組包括多個複製子。多方面的實驗結果表明,大多數生物內DNA的複製都是從固定的起始點以雙向等速方式進行的。複製叉以DNA分子上某一特定順序為起始點,向兩個方向等速生長前進。
(三)DNA複製過程
1.DNA雙螺鏇的解鏇
DNA在複製時,其雙鏈首先解開,形成複製叉,而複製叉的形成則是由多種蛋白質及酶參與的較複雜的複製過程(1)單鏈DNA結合蛋白(single—strandedDNAbindingprotein,ssbDNA蛋白)ssbDNA蛋白是較牢固的結合在單鏈DNA上的蛋白質。原核生物ssbDNA蛋白與DNA結合時表現出協同效應:若第1個ssbDNA蛋白結合到DNA上去能力為1,第2個的結合能力可高達103;真核生物細胞中的ssbDNA蛋白與單鏈DNA結合時則不表現上述效應。ssbDNA蛋白的作用是保證解鏇酶解開的單鏈在複製完成前能保持單鏈結構,它以四聚體的形式存在於複製叉處,待單鏈複製後才脫下來,重新循環。所以,ssbDNA蛋白只保持單鏈的存在,不起解鏇作用。
(2)DNA解鏈酶(DNAhelicase)
DNA解鏈酶能通過水解ATP獲得能量以解開雙鏈DNA。這種解鏈酶分解ATP的活性依賴於單鏈DNA的存在。如果雙鏈DNA中有單鏈末端或切口,則DNA解鏈酶可以首先結合在這一部分,然後逐步向雙鏈方向移動。複製時,大部分DNA解鏇酶可沿滯後模板的5’—〉3’方向並隨著複製叉的前進而移動,只有個別解鏇酶(rep蛋白)是沿著3’—〉5’方向移動的。故推測Rep蛋白和特定DNA解鏈酶是分別在DNA的兩條母鏈上協同作用以解開雙鏈DNA。
(3)DNA解鏈過程
 岡崎片段
岡崎片段2.岡崎片段與半不連續複製
因DNA的兩條鏈是反向平行的,故在複製叉附近解開的DNA鏈,一條是5’—〉3’方向,另一條是3’—〉5’方向,兩個模板極性不同。所有已知DNA聚合酶合成方向均是5’—〉3’方向,不是3’—〉5’方向,因而無法解釋DNA的兩條鏈同時進行複製的問題。為解釋DNA兩條鏈各自模板合成子鏈等速複製現象,日本學者岡崎(Okazaki)等人提出了DNA的半連續複製(semidiscontinuousreplication)模型。
1968年岡崎用3H脫氧胸苷短時間標記大腸桿菌,提取DNA,變性後用超離心方法得到了許多3H標記的,被後人稱作岡崎片段的DNA。延長標記時間後,岡崎片段可轉變為成熟DNA鏈,因此這些片段必然是複製過程中的中間產物。另一個實驗也證明DNA複製過程中首先合成較小的片段,即用DNA連線酶溫度敏感突變株進行試驗,在連線酶不起作用的溫度下,便有大量小DNA片段積累,表明DNA複製過程中至少有一條鏈首先合成較短的片段,然後再由連線酶鏈成大分子DNA。一般說,原核生物的岡崎片段比真核生物的長。深入研究還證明,前導鏈的連續複製和滯後鏈的不連續複製在生物界具有普遍性,故稱為DNA雙螺鏇的半不連續複製。
3.複製的引發和終止
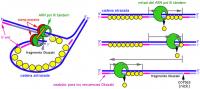
所有的DNA的複製都是從一個固定的起始點開始的,而DNA聚合酶只能延長已存在的DNA鏈,不能從頭合成DNA鏈,新DNA的複製是如何形成的?經大量實驗研究證明,DNA複製時,往往先由RNA聚合酶在DNA模板上合成一段RNA引物,再由聚合酶從RNA引物3’端開始合成新的DNA鏈。對於前導鏈來說,這一引發過程比較簡單,只要有一段RNA引物,DNA聚合酶就能以此為起點,一直合成下去。對於後隨鏈,引發過程較為複雜,需要多種蛋白質和酶參與。後隨鏈的引發過程由引發體來完成。引發體由6種蛋白質構成,預引體或引體前體把這6種蛋白質結合在一起並和引發酶或引物過程酶進一步組裝形成引發體。引發體似火車頭一樣在後隨鏈分叉的方向前進,並在模板上斷斷續續的引發生成滯後鏈的引物RNA短鏈,再由DNA聚合酶III作用合成DNA,直至遇到下一個引物或岡崎片段為止。由RNA酶H降解RNA引物並由DNA聚合酶I將缺口補齊,再由DNA連線酶將每兩個岡崎片段連在一起形成大分子DNA.。
端粒和端粒酶
 岡崎片段
岡崎片段在弄清楚DNA複製過程之後,20世紀70年代科學家對DNA複製時新鏈5’端的RNA引物被切除後,空缺是如何被填補的提出了質疑。如不填補豈不是DNA每複製一次就短一點。以後隨鏈複製為例,當RNA引物被切除後,岡崎片段之間是由DNA聚合酶I催化合成的DNA填補之,然後再由DNA連線酶將它們連線成一條完整的鏈。但是DNA聚合酶I催化合成DNA時,需要自由3’—OH作為引物,最後餘下子鏈的5’無法填補,於是染色體就短了一點。在正常體細胞中普遍存在著染色體酶複製一次端粒就短一次的現象。
人們推測,可能一旦端粒縮短到某一閾限長度一下時,他們就會發出一個警報,指令細胞進入衰老;或許是當細胞判斷出它們的染色體已變得太短了,於是分裂也就停止了,造成正常體細胞壽命有一定界限。但是在癌細胞中染色體端粒卻一直維持在一定長度上,這是為什麼?這是因為DNA複製後,把染色體末端短缺部分補上需要端粒酶,這是一種含有RNA的酶,它既解決了模板,又解決了引物的問題。在生殖細胞和85%癌細胞中都測出了端粒酶具有活性,但是在正常體細胞中卻無活性,20世紀90年代中期,Blackburn首次在原生動物中克隆出端粒酶基因。
端粒酶在癌細胞中具有活性,它不僅使癌細胞可以不斷分裂增生,而且它為癌變前的細胞或已經是癌性的細胞提供了時間,以積累附加的突變,即等於增加它們複製,侵入和最終轉移的能力。同時人們也由此萌生了開發以端粒為靶的藥物,即通過抑制癌細胞中端粒酶活性而達到治療癌症的目的。
至於真核細胞DNA末端的結構特點,早就在1978年Blackburn就以原生動物四膜出(一種纖毛蟲)為例說明之:①迥紋形式的髮夾環;②僅由C,A組成的簡單序列大量重複(C4A2)20~70;③鏈上有許多缺口(nicks)。
