
致死突變型
正文
導致個體死亡的突變型,在高等動植物中常導致個體在性成熟前死亡。致死突變型對於致死的個體來說是不利的,但有利於維持自然群體的雜合狀態。某些致死突變型的生物品系可用於個體發育的研究及育種實踐,所以受到人們的重視。簡史 1905年法國學者L.居埃諾首先報導小鼠的黃色突變型從來沒有育成過純種。這種使小鼠體毛呈黃色的突變基因AY對黑色基因 a雖然是顯性的,但所有黃鼠近交所產生的後代都是2黃色∶1黑色,而不是通常應出現的3∶1分離比(見孟德爾定律)。這種黃色小鼠與黑色小鼠雜交的子代則是1黃色∶1黑色。這表明黃色的小鼠是雜合體。後來的研究證明了含兩個AY基因的純合體黃色小鼠的胚胎曾在母體內出現過,但都在發育早期死亡了。此外又發現將黃色小鼠自交產生的子代個體數同黃色小鼠和黑色小鼠雜交所產生的子代個體數相比平均約少1/4。這些事實證明 AY基因有多效性:它既是一個影響毛色的顯性基因,又是一個影響生活力的隱性致死基因。AY基因純合體的胚泡植入子宮壁後不久便死亡,所以這種小鼠的黃色變種不可能育成純種。上述雜交結果可以表示如下:
親代AYa X AYa
(黃色) ↓ (黃色)
子代 1AYAY : 2AYa : 1aa
(死亡)(黃色) (黑色)
此後在異體交配的動、植物群體及人類中普遍地發現了隱性致死突變型。這種突變型在雜合狀態下一般不影響個體的生活力,但純合狀態則會引起死亡。例如在植物中的隱性白苗致死突變型在純合情況下幼苗缺乏合成葉綠素的能力,子葉中的養料耗盡就會死亡。又如人類中的一種致死性大皰性表皮鬆解症是常染色體隱性遺傳疾病,如果隱性致死基因呈純合狀態則個體在胚胎期死亡。
類別 致死突變型按表現程度可分為全致死的(純合體全部死亡或至少90%死亡),亞致死的(純合體死亡率達50~90%),半致死的(純合體死亡率在10~50%之間)和弱致死的(即類正常的,純合體死亡率在10%以下) 幾種。按環境條件對致死效應的影響又可分為條件致死的(例如溫度敏感突變型在較低溫度下不出現致死效應在較高溫度下出現致死效應)和非條件致死的(在已知任何條件下都有致死效應)兩類。按致死作用發生的階段還可分為配子致死的(花粉死亡)和合子致死的(受精後合子不能發育)、胚胎致死的和幼體致死的。按照個體發育中致死作用時相來分,又可以分為單相致死的(只在某一時期致死)、多相致死的(致死期有兩個或兩個以上)、無相致死的(致死作用可以發生在任何一個時期)。
致死突變可發生在任何染色體上;發生在常染色體上的稱常染色體致死,發生在性染色體上的稱為伴性致死。在果蠅等性染色體屬XY型的生物中,如果隱性致死突變發生在 X染色體上,對雄性果蠅即可產生致死效應;但對雌性果蠅則只有兩個隱性致死突變基因純合才會造成死亡。如果在雌性果蠅的一個 X染色體上帶有致死基因1,在另一個X染色體上帶有它的等位顯性基因 L,雌果蠅就可以生活和產生後代。所以雌性果蠅可以是隱性致死基因的長期攜帶者。這種雌性雜合體(L1)果蠅和正常的雄性果蠅交配所產生的子代的雌雄比不是1♀∶1♂而是2♀∶1♂。
顯性的致死突變型在雜合狀態下即可致死,因此顯性致死突變型只有條件致死的,半致死的或弱致死的。例如人類的Ⅱ型家族性高脂蛋白血症是常染色體顯性突變型,它的主要臨床特徵是高血脂和多發黃色瘤,部分患者常早年死於心肌梗塞。
檢測 觀察雜交的子代是否出現不正常分離比、異常的雌雄比和有規律的早期夭折等都有助於發現致死突變型。1927年美國遺傳學家H.J.馬勒首創 C1B法用來檢測果蠅伴性致死突變基因。這裡1(lethal)代表X染色體上的一個隱性致死基因;B代表同一X染色體上的顯性棒眼基因(bareye,B);C(crossover suppressor)代表 X染色體上一段包括1和B的倒位,它能抑制交換的發生(見染色體畸變),從而保證1和B在連續的世代中穩定地聯繫在一起,所以棒眼性狀的存在就表明這一隱性致死基因的存在。檢測方法是用誘變因子(如 X射線)處理雄性果蠅,然後將它和 C1B雌性果蠅交配,子一代會出現棒眼雌蠅,把子一代中的棒眼雌蠅與野生型雄蠅作單對交配,子二代雌雄蠅數目之比應是2∶1。但是如果在子一代的雌蠅中來自雄性親體的 X染色體上發生了一個隱性致死突變,那么子二代中就看不到雄性果蠅(圖1)。套用這一方法可以檢測 X連鎖隱性致死基因突變率。馬勒用這種方法測得的果蠅 X連鎖隱性致死自發突變率大約是0.1~0.2%。以後又發展起來的Muller-5法、駢連X染色體法等可以用來更有效地檢測果蠅的X連鎖隱性致死突變率。
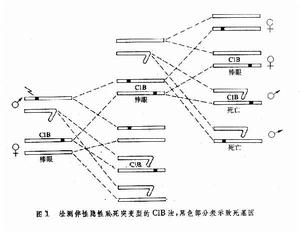 致死突變型
致死突變型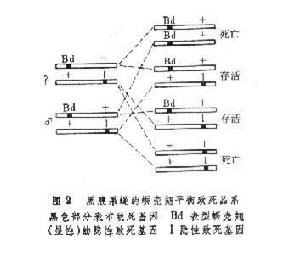 致死突變型
致死突變型致死突變型還可以套用於育種實踐,1975年B.A.斯特魯尼科夫利用性連鎖平衡致死原理育成了幾乎只產雄蠶的家蠶品系。雄性家蠶的性染色體是ZZ,雌性家蠶是ZW。11和12是兩個Z染色體上的致死基因,將不帶有致死基因的雌蛾WZ和兩個 Z染色體上分別帶有致死基因11和12的雄蛾進行雜交,雌蛾所產的蠶卵中雌性蠶卵WZ(11)或WZ(12)全部不能孵化, 而雄性蠶卵++/11+或++/+12則可以孵化(圖3)。
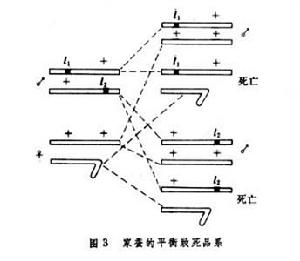 致死突變型
致死突變型