
簡介
研究基因如何控制發育的遺傳學分支學科。從遺傳學觀點看來發育是從基因型轉化為表型的過程。從受精開始,以至胚層、器官原基的形成,組織、細胞的決定和分化,每一步都要受特定基因的控制。這些基因發生突變就會相應地造成發育的異常、停頓、甚至胚胎的死亡。發生遺傳學在方法學上主要是利用這些影響發育的突變型,並結合實驗胚胎學、細胞生物學和分子生物學的方法,從不同水平來分析基因和性狀發育之間的關係,以闡明基因控制發育的機理。因此它同胚胎學、畸胎學、細胞生物學和分子生物學,尤其是與細胞分化過程中基因表達及其調控方面的研究有著密切的關係。
簡史
遺傳是發育的基礎,而發育是遺傳的實現。兩者之間的關係歷來是遺傳學家和胚胎學家共同關心的問題。早在19世紀末期,德國生物學家A.魏斯曼就曾經試圖建立發育和遺傳的統一理論。他曾經假定全部發育過程是受細胞核控制的,卵裂過程中核內遺傳物質的不等分配是胚胎分化的主要原因。這一理論的原有形式雖然被早期實驗胚胎學的實驗事實所否定,但他提出的問題卻吸引了幾代生物學家的繼續探討。
美國細胞學家E.B.威爾遜1928年在《發育和遺傳中的細胞》一書中提出基因在細胞水平的活動是發育的根本原因這一論點,認為發育是“遺傳特性按一定時、空秩序的表現”。基因論的創建人美國遺傳學家兼實驗胚胎學家T.H.摩爾根在他晚年(1934)的著作《胚胎學和遺傳學》一書中也強調遺傳學和胚胎學統一的重要性,並提出在發育的“不同時期有不同的一組基因起作用”的論點。
最早用實驗方法確定了染色體在發育中的重要性的是德國實驗胚胎學家T.H.博韋里。他根據對海膽受精卵分裂球發育的分析結果,認為正常發育依賴全套染色體的正常組合,每一個染色體對發育都有特殊的影響。美國遺傳學家R.B.戈德施米特對基因在發育中的作用給以很大的重視。他在1935年發現擬表型,認為基因突變和環境因子的作用一樣,都可能幹擾相同的發育過程。致死基因和畸型發育的研究對發生遺傳學的形成也起過重要的推動作用。1955年瑞士實驗胚胎學家E.哈多恩在《發生遺傳學和致死因子》一書中系統地總結了致死突變型引起異常發育的大量材料,並用實驗胚胎學方法對這些病理髮育過程進行分析,促進了對正常發育中基因作用機制的了解。1963年H.格呂內貝格在《發育病理學》一書中提出基因作用多效性概念,認為胚胎是一個高度複雜的相互作用系統,任何與發育有關的突變都可能對胚胎髮育產生廣泛的影響。因此應透過對各種病理髮育現象的分析找出基因在細胞水平作用的原初效應。70年代初 C.L.馬克特和 H.烏爾施普龍合寫了《發生遺傳學》,把當時散見於各方面的有關資料匯總起來,初創了這門學科的體系。
長期以來胚胎學家致力於用實驗方法分析發育的生理原因,尤其著重組織者的研究,因而忽視了發育的遺傳基礎。對異屬誘導系統的實驗結果說明誘導者只起激發作用,被誘導出來的器官的種屬特性則取決於起反應的細胞本身的遺傳特性。例如把蛙的外胚層移植到蠑螈的頭部,被誘導形成的口器是蛙特有的角質齶和吸盤。這些事實促使胚胎學家注意發育和遺傳關係問題。1940年英國實驗胚胎學家兼遺傳學家C.H.沃丁頓在《組織者和基因》一書中首先提出從基因和細胞質環境的相互作用來理解反應能力、誘導和決定等胚胎學基本概念的主張。但限於當時生物學發展水平,還不能對基因控制發育的分子機製作深入的研究。
50年代中期以來的分子生物學的重大進展使解決遺傳和發育關係問題的條件逐漸成熟起來。遺傳信息傳遞的中心法則揭示了生物的遺傳和發育的內在聯繫。從分子水平看來,細胞分化和性狀發育都是表型專一的大分子合成的結果,因而歸根結蒂依賴基因在發育過程中按一定的時空秩序的表達。基因的表達又可以用相應的信使核糖核酸(mRNA)的轉錄和專一的蛋白質(如結構蛋白、酶等)的合成來追蹤。1976年美國分子生物學家E.戴維森所寫的《早期發育中的基因活動》一書代表從分子水平探討發育和遺傳關係問題的發展趨向。目前研究核酸分子結構和功能的方法日臻完善,尤其是遺傳工程技術提供了前所未有的研究基因結構、表達及其調控的有力的手段。這就為從分子水平探討發生遺傳學問題,特別是研究影響特定發育過程的單個基因(或基因群)的作用開闢了新的前景。
對象和方法
發生遺傳學開始於對黑腹果蠅發育的遺傳學分析。從摩爾根時代以來已積累了大量有關果蠅遺傳和發育的實驗資料和許多影響發育的突變型。果蠅的唾腺巨大染色體上疏鬆區的消長可以反映不同發育時期的基因活動情況。此外,對體表形態構造的變異也便於作精細的描述。這些都是果蠅這一實驗材料的優點,缺點是較難得到足夠量的可供生化分析的樣品。另一個常用的材料是小鼠,也有許多遺傳背景清楚的純系和致死突變型(如T基因座位)可供發生遺傳學研究,不過同樣存在取材上的困難。用海膽作為研究對象則便於取得較大量的材料,特別是同一發育階段的材料,可是可供研究的突變型較少。英國分子生物學家 S.布倫納在 60年代倡導用一種營自由生活而繁殖迅速的秀麗隱桿線蟲(Caenorabditis elegans) 作材料。這種動物身體和各器官的細胞數少而恆定,胚胎髮育屬於鑲嵌型,每一個器官的來源有確定的細胞譜系可循,而且便於用實驗方法得到許多突變型。低等真核生物如粘菌(Dictyosteliumdiscoideum)由於生活史和形態發生的特點,也被一些人用來作為發生遺傳學的研究對象。此外,甚至有人採用更為簡單的對象如細菌形成芽孢的過程、噬菌體的自動裝配過程等作為一個發育模型來進行發生遺傳學研究。
套用突變型進行遺傳學分析是發生遺傳學的基本研究方法。此外,實驗胚胎學方法(如移植、離體培養等)、細胞生物學方法(如核移植、細胞化學技術等)、生化方法 (如同功酶測定等)和分子生物學方法(如重組DNA技術、分子雜交等)都是常用的實驗手段。70年代以來重組DNA技術的套用使有可能直接研究沒有發生突變的野生型基因的作用。
研究主題
動物的發育是從受精開始,通過受精卵的核質之間、分裂球之間、以及胚胎不同部位之間的相互作用,使基因按一定時空秩序表達,從而控制細胞和器官原基的逐步決定和分化的過程。動物的發育類型(鑲嵌卵或調整卵)不同,基因控制發育的具體方式也可能不同。果蠅(鑲嵌卵)的發育中細胞的發育命運決定得早,胚胎髮育主要表現為一連串由大到小的發育區逐步劃分的過程。兩棲類(調整卵)的發育中細胞的命運決定得晚,細胞遷移和相互作用在發育中起重要的作用。截至20世紀70年代發生遺傳學的知識還主要是從果蠅上得到的。
核質關係和母體效應
細胞核移植實驗證明果蠅前囊胚層細胞核具有全部發育潛能,而囊胚層細胞核的發育潛能已開始受到限制,這可能是受到卵表層細胞質的影響的結果。紫外線損傷和卵質移植實驗證明位於卵後端的極質能決定遷移到這區域中的細胞核的發育命運,使它們形成原始生殖細胞。無尾兩棲類(如蛙和瓜蟾)中也存在類似的現象,它們的原始生殖細胞是由位於卵的植物極的生殖質決定的。
絕孫突變型雌性果蠅的囊胚細胞核不能及時遷入極質區域,生殖細胞因而不能正常發生。這表明卵質特性是在卵子發生過程中受母體基因決定的,這一現象稱為母體效應。某些母體效應突變型如雙腹端(bicaudal,bic)和背方 (dorsal,dl)能廣泛影響胚胎髮育的格局。純合雙腹端 (bic/bic)雌蠅所產的一部分卵只發生成為兩個腹端,彼此沿前後軸對稱排列,而缺少頭、胸和其他腹節。純合背方 (dl/dl)雌蠅的胚胎構造全部背方化,而前後極性仍然保持。這兩個母體效應基因各自定位在染色體的一個位置上。有人假定它們的野生型基因產物的濃度在卵內各自沿前後軸和背腹軸呈梯度分布,從而決定了果蠅胚胎的正常發育格局。
細胞的決定和發育區劃分
利用帶有不同遺傳標記的胚胎細胞的嵌合體可以追溯細胞決定的時期、發育命運以及器官原基奠基細胞的數目。
得到嵌合體的常用方法有3種:
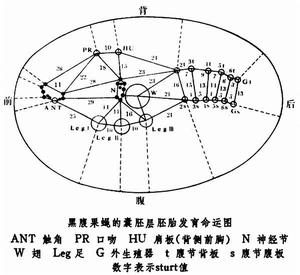
①雌雄嵌合體,在果蠅中可以利用環狀X染色體常在受精卵的第一次有絲分裂過程中丟失的特性取得雌雄嵌合體。例如雌性親本的兩個 X染色體上全部都是野生型基因,不過其中一個是環狀染色體;雄性親本的一個X染色體上帶有3個隱性突變基因(白眼w,黃體y和分叉剛毛bi)。這兩個親本交配後可以產生一種帶有一個環狀X染色體和一個有 3個隱性基因的X染色體的受精卵。這種受精卵第一次卵裂核分裂後,一個子細胞帶有這兩個X染色體,因而發育成嵌合體上具有野生型表型的雌性部分(XX);另一個子細胞由於丟失了環狀 X染色體,因而發育成為具有突變型表型的雄性部分(X0)。兩部分之間有清楚的分界面。由於第一次核分裂的紡錘體的取向不同,含XX和X0細胞群的分界面也有不同。因此所得到雌雄嵌合體可以是左右各半、前後各半或者是其他種種形式。利用雌雄嵌合體的這些特點,便可以製作各器官原基在囊胚層上的預定命運圖。其原理是兩個成體器官(如翅和頭)的原基細胞在囊胚層上相距愈近,則嵌合體分界線通過兩原基之間的機率就愈小。因此帶有不同遺傳標記的兩個構造在嵌合體上出現的百分比可作為這兩個原基之間相對距離的量度(為了紀念首先提出這種構想的A.H.斯特蒂文特,用sturt作為單位,一個sturt代表 1%個體中嵌合體分界線通過所研究的兩個原基)。如果再測出這兩個構造中每一個對第三個構造(如吻)的相對距離就可以確定某一器官的原基在囊胚層上的相對位置。推而廣之,便可以做出各器官在囊胚層上的預定命運圖(見圖)。
 ②重組嵌合體,在一定發育時期用X射線誘發胚胎細胞中的有絲分裂交換(見連鎖和交換),使單個胚胎細胞及其子細胞群帶上遺傳標記,便可鑑定發育時期細胞的決定狀態。例如用X射線照射小體(M)雜合體(M/M+)果蠅的囊胚層期胚胎,可以誘發體細胞染色體交換而在生長緩慢的雜合體細胞背景上出現一片生長迅速的野生型純合M+/M+細胞。這種細胞群決不跨越兩個體節,說明體節的決定可能發生在囊胚層期或稍後的一次分裂期。晚一些時候照射所得到的一個標記細胞群決不越過同一體節的前部和後部的分界線,說明體節的前部和後部這時已經決定。再晚些時候照射,得到的一個標記細胞群就不再同時包含背方和腹方的構造,也就是說發生了背腹的分區。器官發生過程中還會逐漸發生更進一步的分離。象這樣每一來源於少數幾個奠基細胞的細胞群所占據身體或器官上的一定區域稱為發育區。屬於一個發育區的細胞決不會越界同其他發育區的細胞混合。發育區可看作是發育的基本單位,它在許多方面具有“場區”的性質。同一發育區內發育格局可以調整,而不同發育區之間則是鑲嵌的、不可調整的。果蠅的發育過程是一系列由大到小發育區愈分愈細的過程。這一現象稱為發育區劃分。
②重組嵌合體,在一定發育時期用X射線誘發胚胎細胞中的有絲分裂交換(見連鎖和交換),使單個胚胎細胞及其子細胞群帶上遺傳標記,便可鑑定發育時期細胞的決定狀態。例如用X射線照射小體(M)雜合體(M/M+)果蠅的囊胚層期胚胎,可以誘發體細胞染色體交換而在生長緩慢的雜合體細胞背景上出現一片生長迅速的野生型純合M+/M+細胞。這種細胞群決不跨越兩個體節,說明體節的決定可能發生在囊胚層期或稍後的一次分裂期。晚一些時候照射所得到的一個標記細胞群決不越過同一體節的前部和後部的分界線,說明體節的前部和後部這時已經決定。再晚些時候照射,得到的一個標記細胞群就不再同時包含背方和腹方的構造,也就是說發生了背腹的分區。器官發生過程中還會逐漸發生更進一步的分離。象這樣每一來源於少數幾個奠基細胞的細胞群所占據身體或器官上的一定區域稱為發育區。屬於一個發育區的細胞決不會越界同其他發育區的細胞混合。發育區可看作是發育的基本單位,它在許多方面具有“場區”的性質。同一發育區內發育格局可以調整,而不同發育區之間則是鑲嵌的、不可調整的。果蠅的發育過程是一系列由大到小發育區愈分愈細的過程。這一現象稱為發育區劃分。
③異表型嵌合體,把基因型不同的兩種小鼠的早期胚胎在體外人工地併合在一起,然後移回到母鼠子宮內,便能發育成嵌合體,稱為異表型嵌合體。通過異表型嵌合體研究,在小鼠中測定了色素細胞、毛囊、生殖腺以及其他內臟的奠基細胞的數目。例如生殖腺的奠基細胞數是2~9。
發育途徑的決定和轉變
果蠅成蟲的器官原基──成蟲盤在胚胎髮育的早期已經決定了,但要等到幼蟲變態時受到激素的影響才開始分化。E.哈多恩曾把成蟲盤(如生殖板原基)移植到成蟲腹腔內,經過連續70次以上的傳代(超過1000次細胞分裂)後再移植到將要變態的幼蟲體內仍然可以按照預定的命運分化為生殖板。這說明細胞的決定和分化在時間上是可以分隔的。細胞雖然經過了上千次的分裂仍然可以保持原來的決定狀態。然而成蟲盤經過若干次移植傳代後偶爾可能改變其決定狀態,分化成和它原來的預定命運不同的器官(如觸角)。這一現象稱為轉決,它說明細胞的發育命運可以改變。
影響果蠅發育途徑的還有一種突變型稱為同源異形突變型,它能使一種發育途徑轉變到另一種發育途徑,例如使平衡棒原基發育成為翅,觸角原基發育成為足等。同源轉化的影響限於一個發育區內,如反雙胸突變(CBX)能將中胸後部(翅的後部)轉化為後胸後部(平衡棒的後部)。根據同源異形突變型的作用方式,有人構想它的野生型基因在正常發育過程中可能起選擇和維持特定發育途徑的作用,因此稱之為選擇基因。果蠅胚胎髮育中發育區的劃分是由大到小逐步進行的,其中每一步都可能受一個(或一組)選擇基因的控制。
發育程式的遺傳控制
在果蠅和小鼠等動物中都發現有一類控制某些酶活性在胚胎髮育中出現的時間和地位的基因,稱為時序基因。例如黑腹果蠅的澱粉酶為2號染色體上兩個頭尾相連的結構基因所編碼。在它們的附近有一個基因map,它的3個等位基因mapa、mapb、mapc控制著澱粉酶在中腸出現的時間和地位。小鼠的 5號染色體上有一個編碼β-葡萄糖苷酸酶的結構基因Gus,和它緊密連鎖的有一個控制酶出現時間的時序基因Gut。Gut基因發生突變後,β-葡萄糖苷酸酶在各種組織中出現的時間便被擾亂。
此外,果蠅等雙翅目昆蟲的幼蟲唾腺染色體上的疏鬆區也是研究基因活動的時空秩序的好材料。疏鬆區是光學顯微鏡下可觀察到的染色體的局部膨大的部分,累積大量新合成的mRNA,是進行活躍的轉錄活動的基因所在的地方。果蠅發育過程中染色體不同部位的疏鬆區按一定的時間順序出現,說明這些基因在不同的發育時期起作用。蛻皮激素能誘發特定的疏鬆區出現,說明這一激素是通過調節該基因的活動而參與控制發育的。
致死突變型也常用作研究發育程式的遺傳控制的材料,因為使胚胎髮育在特定階段停頓的基因中間有許多是控制該特定發育階段的基因。許多這類突變型是溫度敏感的,在果蠅和線蟲都獲得了一些溫度敏感的突變型,而且已發現不同的突變型常使發育停頓在不同的階段,某些突變型還影響幾個發育階段。
基因影響發育的機制
影響發育的基因究竟通過什麼途徑或基因產物起作用,這也是必須回答的問題。這方面研究得較為深入的是小鼠的T複合座位或T基因複合體,它位於17號染色體上,包括6個互補群。
除野生型基因和顯性的突變型T以外,還包括一系列的隱性致死或半致死突變型t。雜合體(T/t)具有短尾,純合體(T/T)則造成胚胎早期畸形和夭折。一系列的隱性致死或半致死突變型 (t)各自對早期發育的一定階段專一地起作用。例如t12影響滋養外胚層和內胚層細胞團的分離而使發育停頓在桑椹期;t0作用在胚外外胚層和外中胚層分離的時期;t9影響原條期而使中胚層細胞向內遷移受阻,因而造成中軸器官發育異常。進一步分析發現基因的原初效應可能影響細胞表面抗原和它的下方的微絲,從而干擾了原腸運動正常進行。
T 基因座位的各個野生型基因的功能可能是控制早期發育中細胞表面特定分子按順序的表達,從而保證形態發生運動和細胞間相互作用按正常的發育程式有條不紊地進行。對於多數控制發育的基因來說,它們究竟編碼什麼蛋白質還不清楚,那些控制基因活動的時空秩序的基因的作用機制則更不了解了。
從分子水平研究基因如何控制發育,對於解決發育和遺傳關係這一生物學基本問題有重要的理論意義。在實踐上,這方面研究的進展又是了解病理髮育(畸胎、腫瘤等)的發生機制,以及把遺傳工程方法套用到高等動物,治療遺傳病或改變經濟動物遺傳性的必要的理論前提。
參考書目
A.D.Stewart & D. M. Hunt,The Genetic Basis of Development,Blackie,1982.
