
發生過程
過程
全部精子發生過程可以被分為 3 個過程
精原細胞位於 生精上皮的基底部,分為 A 、 B 兩種類型。 A 型 精原細胞進一步分為 Ad 型和 Ap 型精原細胞。在正常情況下, Ad 型 精原細胞不發生任何有絲分裂,應該被視為精子發生的 精原幹細胞; Ap 型精原細胞則通常 分化 增殖為兩個 B 型精原細胞。 B 型 精原細胞分裂增殖為 初級精母細胞,隨後,初級精母細胞開始 DNA 合成過程。
精母細胞經歷了 減數分裂的不同階段。 粗線期時 RNA 的合成十分活躍。 減數分裂的結果產生單倍體生精細胞,又稱 精子細胞。在精子生髮過程中, 減數分裂是一個非常關鍵的過程,在這個階段, 遺傳物質相互重組、染色體數目減少並最終形成 精子細胞。次級精母細胞產生於第一次 減數分裂後。這些生精細胞含有雙份單倍體染色體。在第二次 減數分裂精母細胞演變為單倍體的 精子細胞。第一次 減數分裂前期大概持續 1 ~ 3 周,而除此之外的第一次 減數分裂的其他階段和第二次減數分裂在 1 ~ 2 天之內完成。
第二次減數分裂後形成 精子細胞,是沒有減數分裂活性的圓形細胞。圓形的 精子細胞經過複雜的顯著變化轉變為不同長度的精子細胞和精子。在第二次 減數分裂中,細胞核發生的聚縮和塑性,同時鞭毛形成和胞漿明顯擴張。全部 精子細胞變形的過程稱為精子形成。
動力學
精子生成過程在時間和空間上有嚴格的順序性。曲細精管中不同 成熟階段的生精細胞在管腔中連續、依次排列,提示精子生成過程按照順序依次從第一階段到第二階段,再到第三、第四階段。這個順序稱為生精波( spermatogenic wave )。在人類以及某些猴的部分睪丸組織中,曲細精管的同一轉化部位也可以同時存在多個精子 成熟階段。曲細精管同一局部的精子生成過程呈螺鏇樣相互聯繫。
除了精子發生的時間和各個階段的空間分布特徵之外,人類的精子發生還表現出另外一個特徵:人類的生精細胞數目相對較少,同樣,每個 支持細胞相對應的生精細胞數目也較低。上述因素以及精子與 支持細胞較低的比率( 1 : 5 )導致人類的精子生成數目相對較少。人類每克睪丸組織在 24 小時內生成的精子數目 300 ~ 700 萬。
生精細胞的 增殖和 分化過程都遵循一個嚴格的模式,所有生精細胞的發育和分化都經過幾個獨立而又緊密聯繫的過程。 生精上皮按照程式成功完成每一個發育階段稱為生精上皮周期,每個周期需要 16 天,人類的精子發育到成熟必須經過四個周期,可以推測起碼需要 64 天。
精子發育成熟釋放到曲細精管管腔的過程稱為精子釋放,而這些過程受到多種因素的影響,包括 血纖維蛋白溶酶原、 激素、溫度、毒性物質。未釋放的精子將被 支持細胞吞噬。
分泌調控
睪丸的生精及合成雄激素兩項功能都通過負反饋受到下丘腦和腦垂體的調節。
睪酮可以抑制LH、FSH的分泌。對於 FSH ,抑制素 B 是更為重要的調節物質。 LH 促進睪丸間質細胞合成睪酮, FSH 則控制支持細胞的調節精子生成作用。睪酮在睪丸間質中的作用對於精子發生過程也十分重要。
精子發生的初次生精過程一般在 FSH 和 LH 的影響下完成。但是高濃度的睪酮單一作用也可以誘導精子發生。在睪酮分泌型睪丸間質細胞瘤的腫瘤附近和 LH 受體激活性突變的患者體內,都可以見到完整的精子發生過程。非常關鍵的治療目的就是試圖在睪丸間質中聚集高濃度的睪酮。臨床常用的辦法是使用hCG,它具有較高的 LH 和 FSH 活性。激素在生精維持、生精再激活中同樣有重要作用。大劑量睪酮通過負反饋機制抑制促性腺激素的分泌,並導致射精中的精子數量大量減少;即使使用 FSH 後精子生成數量也只能達到正常數量的 30% 。與之相似,使用 hCG 後也可以造成生精數量減少,其機制是由於 hCG 刺激產生的睪酮發揮了負反饋抑制,但是其抑制生精的作用不如單獨使用睪酮的效果明顯。而且, hCG 的生精抑制作用可以在使用 FSH 後完全恢復。 hCG 和睪酮抑制生精的效果差異是由於在睪丸間質中睪酮的濃度更高。
使用抗體免疫中和 FSH 可以明顯減少靈長類動物以及男子人類的精子發生。在抑制內生性促性腺激素分泌後, FSH 可以持續地維持生精過程。最近的證據發現在腦垂體切除的患者中,在缺少 LH 、 FSH 受體激活性突變的情況下,生精功能可以正常存在。儘管還不知道睪丸間質的睪酮濃度,但是這例患者提示 FSH 受體結構激活對於正常生精是十分必要的。推測睪酮的作用可能是激活 FSH 受體,使 FSH 與其結合後發揮作用。
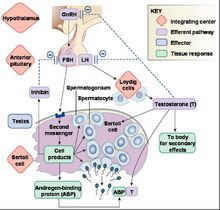 精子發生分泌調控流程圖
精子發生分泌調控流程圖LH 、 FSH 以及睪酮的協同作用對維持正常生精和生精再激活必不可少。
基因調控
精子發生期間染色質濃縮,使 DNA不能夠轉錄,這種情況在精子完全形成之前完成。各種動物在精子形成中轉錄停止的時刻不完全相同。例如在果蠅,RNA合成在 初級精母細胞期間停止,而在小鼠,在成熟分裂後不久的 精子細胞中還在進行,要在細胞核開始伸長時才完全停止。
正常精子
精子的形成依賴蛋白質合成,既然 RNA合成已停止,精子變態所需蛋白質合成必然要依賴較早時期產生並儲藏起來的到精子變態時才進行轉譯的穩定RNA,這是發生在轉錄後水平上的調控,是延遲基因表達的機制。如在 精子細胞質中合成並進入細胞核取代組蛋白的魚精蛋白,其基因在 初級精母細胞中就已轉錄。在核內合成的 RNA轉移到細胞質中,與蛋白質結合形成 16~18S的核蛋白顆粒,並以這種形式儲藏在細胞質內直到 精子細胞時期。在這種轉錄和轉譯之間有較長時間間隔的例子中,對控制轉錄後基因表達的因素尚缺乏了解。類似的現象可能也會在其他類型細胞的終末 分化中遇到。
