
特徵描述
種群密度
主條目:種群密度
種群密度是指在單位面積或體積中的個體數,“種群密度”與“密度”不同,前者是個體的“數目”,後者是比例,種群密度是種群最基本的數量特徵。農林害蟲的預報、漁業上捕撈強度的確定等,都需要對種群密度進行調查。自然狀態下一個種群的種群密度往往有著很大的起伏,但不是無限制的變化。出生率、死亡率、遷入與遷出率對種群密度都有影響。種群的大小有上限和下限。種群密度的上限由種群所處生態系統的能量流動決定,下限不好確定,生態系統的穩態調節可以使優勢生物的種群密度保持在一個有限的範圍內 。
種群密度的統計與估算方法
種群密度在生產生活中有重要作用,以下介紹兩種常用的統計與估算方法,估算時“等可能”最為關鍵,不能摻入人為因素。
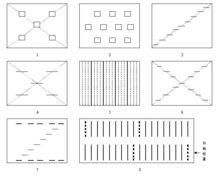 常見的取樣方法
常見的取樣方法①樣方法
樣方法適合調查植物,以及活動能力不強的動物,例如,跳蝻、蝸牛,蒲公英等。其操作過程是:在被調查範圍內,隨機選取若干個完全相等的樣方,統計每個樣方的個體數,並求出每個樣方的種群密度,再求出所有樣方種群密度的均值,以此值作為被調查種群之種群密度的估算值。
常見的取樣方法有“等距取樣法”、“五點取樣法”、“Z字取樣法”等。
②標誌重捕法
標誌重捕法適用於活動能力較強,活動範圍較大的動物,例如,田鼠、鳥類、魚類等。其操作過程是:在被調查種群的活動範圍內,捕獲部分個體,作上標記,再放回原來的環境中,經過一段時間後在同一地點進行重捕,估算公式:
種群總數/標記個體數=重捕個數/重捕中標記個體數
此估算方法得出的估算值傾向於偏大,因為很多動物在被捕獲一次後會更加難以捕獲,導致“重捕中標記個體數”偏小。標記時也需要注意,所用標誌要小而輕,不能影響生物行動;也不能用過於醒目的顏色(比如紅色),否則會使生物更加容易被天敵捕食,影響估算精確度。
出生率與死亡率
出生率指在一特定時間內,一種群新誕生個體占種群現存個體總數的比例;死亡率則是在一特定時間內,一種群死亡個體數占現存個體總數的比例。自然狀態下,出生率與死亡率決定種群密度的變化。出生率大於死亡率,種群密度增長,其他情況同理。
遷入率與遷出率
許多生物種群存在著遷入、遷出的現象,大量個體的遷入或遷出會對種群密產生顯著影響。對於一個確定的種群,單位時間內遷入或遷出種群的個體數占種群個體總數的比例,分別成為種群的遷入率和( immigration rate)遷出率( emigration rate)。遷入與遷出率在現代生態學對城市人口的研究中占有重要地位。
性別比例
性別比例是指種群中雌雄個體的數目比,自然界中,不同種群的正常性別比例有很大差異,性別比例對種群數量有一定影響,例如用性誘劑大量誘殺害蟲的雄性個體,會使許多雌性害蟲無法完成交配,導致種群密度下降.
年齡結構
種群的年齡結構是指一個種群幼年個體(生殖前期)、成年個體(生殖時期)、老年個體(生殖後期)的個體數目,分析一個種群的年齡結構可以間接判定出該種群的發展趨勢。
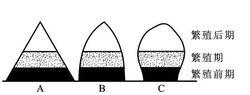 年齡結構,增長型(A)、穩定型(B)
年齡結構,增長型(A)、穩定型(B)增長型
在增長型種群中,老年個體數目少,年幼個體數目多,在圖像上呈金字塔型,今後種群密度將不斷增長,種內個體越來越多。
穩定型
現階段大部分種群是穩定型種群,穩定型種群中各年齡結構適中,在一定時間內新出生個體與死亡個體數量相當,種群密度保持相對穩定。
衰退型
衰退型種群多見於瀕危物種,此類種群幼年個體數目少,老年個體數目多,死亡率大於出生率,這種情況往往導致惡性循環,種群最終滅絕,但也不排除生存環境突然好轉、大量新個體遷入或人工繁殖等一些根本扭轉發展趨勢的情況。
空間格局
組成種群的個體在其空間中的位置狀態或布局,稱為種群空間格局。種群的空間格局大致可分為3類:
 均勻型
均勻型均勻分布uniform) 均勻型分布,指種群在空間按一定間距均勻分布產生的空間格局。根本原因是在種內鬥爭與最大限度利用資源間的平衡。很多種群的均勻型分布是人為所致,例如,在農田生態系統中,水稻的均勻分布。自然界中亦有均勻型分布,例如,森林中某些喬木的均勻分布。
 隨機型
隨機型隨機分布(random)
隨機型分布,是指中每一個體在種群領域中各個點上出現的機會是相等的,並且某一個體的存在不影響其他個體的分布。隨機分布比較少見,因為在環境資源分布均勻,種群內個體間沒有彼此吸引或排斥的情況下,才易產生隨機分布。例如,森林地被層中的一些蜘蛛,麵粉中的黃粉蟲等。
 成群型
成群型集群分布 clumped) 成群分布是最常見的內分布型。成群分布形成的原因是:
⑴環境資源分布不均勻富饒與貧乏相嵌;
⑵植物傳播種子方式使其以母株為擴散中心;
⑶動物的社會行為使其結合成群。
成群分布又可進一步按群本身的分布狀況劃分為均勻群、隨機群和成群群,後者具有兩級的成群分布。
動態變化
數學模型
種群中一些簡單的、具有典型性的動態變化可以用數學模型衡量,常見的有兩種
指數增長("J"型增長)
指數增長模型的提出者是著名人口學家托馬斯·馬爾薩斯( T.Maithus),他認為種群數量的增長不是簡單的相加關係,而是成倍地增長;後來,生物學家查爾斯·羅伯特·達爾文( C.R.Darwin)通過對大象種群的研究再次確認了這一增長模式 。這種客觀存在的增長模式表明,所有種群都有爆炸式增長的能力 。
指數增長的函式式是指數方程,變數為時間t,常數為種群密度增長的倍數。這一增長模式沒有上限,完全的指數增長只存在於沒有天敵、食物與空間絕對充足(以至於沒有種內鬥爭)的理想情況,實際生活中,培養皿中剛接種的細菌、入侵生物(例如鳳眼蓮)、藍藻爆發時,種群會在相當一段時間內進行指數增長,隨後則趨於穩定或大量死亡。
邏輯斯蒂增長("S"型增長)
主條目:邏輯斯蒂增長模型
指數增長是一種過於理想的情況,許多生物在指數增長一段時間後,數量會維持穩定,這可以用另一個數學模型進行描述。
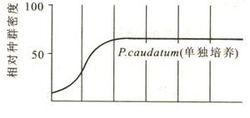 試驗結果
試驗結果實例:俄羅斯生態學家G.W.高斯( G.W.Gaose)曾進行試驗,在0.5ml培養液中放入5個大草履蟲,每24h統計一次該種群的種群密度,結果見右圖,由圖可知,大草履蟲在進行了快速的增長後,穩定在75隻(K值)這個數量上。
邏輯斯蒂增長模型能更好地指導人為的種群調節。
環境容納量( carrying capacity)
進行邏輯斯蒂增長的種群在數量上,存在一個上限,這個上限就被稱為環境容納量,簡記“K值”,代表在環境在不受到破壞的情況下對該種群最大承載量,或該種群在該環境的最大數量。一個種群在種群密度為K/2時,增長率最快,這可以指導經濟生物的採集,讓種群密度始終控制在K/2的範圍內,“多餘”的進行採集,可以讓經濟生物保持最快的增長。
自然增長與下降
自然界中,一個種群的數量變化並不是只增不減,也未必完全符合上述數學模型,其數量變化有一些基本特性
周期性變化
①季節性變化
一般具有季節性生殖的種類,種群密度的最大值常落在一年中最後一次繁殖之後,以後繁殖停止,種群因只有死亡而無生殖,故種群密度下降,這種下降一直持續到下一年繁殖季節的開始,這時是種群數量最低的時期,由此出現季節性的變化。
實例:在歐亞大陸寒帶地區,許多小型鳥獸,通常由於冬季停止繁殖,到春季開始繁殖前,其種群數量最低。到春季開始繁殖後數量一直上升,到秋季因寒冷而停止繁殖以前,其種群數量達到一年的最高峰。體型較大,一年只繁殖一次的動物,如狗獾,旱獺等,其繁殖期在春季,產仔後數量達到高峰,以後由於死亡,數量逐漸降低。
對種群密度有季節性變化的種群做調查時,通常要進行兩次。
②年變化
在環境相對穩定的條件下,種子植物及大型脊椎動物的種群密度在較長的時間跨度內呈現周期性變化。例如:常見的喬木如楊、柳每年開花結果一次,其種子數量相對穩定;又如大型有蹄類動物,一般每年產仔1~2個,其種群數量相對穩定。加拿大盤羊36年的種群數量變動,其最高與最低量的比率僅為4.5倍。而美洲赤鹿在20餘年冬季數量統計中,其最高量與最低量之比只有1.8倍。
不規則波動
動物中還有一些數量波動很劇烈,但不呈周期性的種類,人們最熟知的是小家鼠。它生活在住宅、農田和打穀場中,據中國科學院的16年統計資料,其年均捕獲率波動於0.10~17.57之間,即最高—最低比率為幾百倍。又如布氏田鼠也具有不規律的數量變動。其數量最低的年代,平均每公頃只有1.3隻鼠,而在數量最高的年份,每公頃可達786隻鼠,兩者竟差600多倍種群中有出生和死亡,其成員在不斷更新之中,但是這種變動都往往圍繞著一個平均密度。即種群受某種干擾而發生數量的上升或下降,有重新回到原水平的傾向。這種情況就是動態平衡。
種群的暴發
具不規則或周期性波動的生物都可能出現種群的暴發,赤潮便是此類情況的實例
種群調節
主條目:種群自然調節
在自然界中,絕大部分種群處於一個相對穩定狀態。由於各種因素的作用,種群在生物群落中,與其他生物成比例地維持在某一特定密度水平上,這被稱為種群的自然平衡,而這個密度水平則叫做平衡密度。
由於各種因素對自然種群的制約,種群不可能無限制的增長,最終趨向於相對平衡,而密度因素是調節其平衡的重要因素。種群離開其平衡密度後又返回到這一平衡密度的過程稱為種群調節。能使種群回到原來平衡密度的因素稱為調節因素。
世界上的生物種群大多已達到平衡的穩定期。這種平衡是動態的平衡。一方面,許多物理的和生物的因素都能影響種群的出生率和死亡率,另一方面,種群有自我調節的能力,通過調節而使種群保持平衡。
密度制約因素
影響種群個體數量的因素很多。有些因素的作用是隨種群密度而變化的,這種因素稱為密度制約因素。
例如,傳染病在密度大的種群中更容易傳播,因而對種群數量的影響就大,反之,在密度小的種群中影響就小;又如,在密度大的種群中競爭強度比較大,對種群數量的影響也較大,反之就較小。
密度制約因素的反饋調節:生物種群的相對穩定和有規則的波動和密度制約因素的作用有關。當種群數量的增長超過環境的負載能力時,密度制約因素對種群的作用增強,使死亡率增加,而把種群數量壓到滿載量以下。當種群數量在負載能力以下時,密度制約因素作用減弱,而使種群數量增長。現舉幾例說明這種反饋調節。
①食物
以生活在加拿大的猞猁為例進行說明,研究人員在90年的時間裡,對猞猁和雪兔的數量進行了研究,發現,猞猁種群密度的變化近似隨雪兔種群密度的變化而變化,雪兔數量多時,猞猁的食物增多,種群密度上升,其它情況同理,這就是食物對種群調節的影響。
②生殖力
生殖力也受密度的影響,池塘內的椎實螺在低密度時產卵多,高密度時產卵就少。英國林區的大山雀,每窩產卵數隨種群密度的大小而減少或增多。但這個效果也可能是由於密度高時食物缺少或某些其他因素的作用所引起的。
③抑制物的分泌
多種生物有分泌抑制物來調節種群密度的能力。在植物中,桉樹有自毒現象,密度高時能自行減少其數量。細菌也有類似的情況:繁殖過多時它們的代謝物就將限制數量的再增加;密度降低時,這些代謝產物少,就不足以起抑制作用,因而數量又能上升。
④疾病、寄生物
種群密度越高,流行性傳染病、寄生蟲病越容易蔓延,結果個體死亡多,種群密度降低。種群密度低了,疾病反而不容易傳染了,結果種群密度逐漸恢復。
非密度制約因素
有些因素雖對種群數量起限制作用,但作用強度和種群密度無關。氣候因素就是這樣,颳風、下雨、降雪、氣溫都會對種群的數量產生影響,但這種因素起多大作用與種群密度也是無關的,這類因素稱為非密度制約因素。
作用:生物種群數量的不規則的變動往往同非密度制約因素有關。非密度制約因素對種群數量的作用一般總是很猛烈的,災難性的。例如,我國歷史上屢有記載的蝗災是由東亞飛蝗( Locusta migra-toria manilensis)引起的。引起蝗蟲大發生的一個物理因素是乾旱。東亞飛蝗在禾本科植物的荒草地中產卵,如果雨水多,蟲卵或因水淹或因黴菌感染而大量死亡,因而不能成災,只有氣候乾旱蝗蟲才能大發生,所以我國歷史上連年乾旱常同時伴隨蟲災。
二者的關係
密度制約因素與非密度制約因素何者對種群密度的影響更大需要具體問題具體分析,物理因素等非密度制約因素雖然沒有反饋作用,但它們的作用可以為密度制約因素所調節,即可以通過密度制約因素的反饋機制來調節的。當某些物理因素髮生巨大變化(如大旱、大寒)或因人的活動(如使用殺蟲劑)而使種群死亡率增加,種群數量大幅度下降時,密度制約因素如食物因素就不再起控制作用,因而出生率就得以上升,而種群數量很快就可恢復到原來的水平。
研究生物種群數量變動的規律和影響數量變動的因素,特別是種群數量的自我調節能力,就有可能制定控制種群數量的措施,對種群數量變動進行預測預報,為生產服務(如制定防治害蟲的規劃,對害蟲、害獸發生的測報,以及決定狩獵與採伐的合理度等。
種內關係
集群
主條目:集群
集群( aggregahon或 society、 colony)現象普遍存在於自然種群當中。同一種生物的不同個體,或多或少都會在一定的時期內生活在一起,從而保證種群的生存和正常繁殖,因此集群是一種重要的適應性特徵。在一個種群當中,一些個體可能生活在一起而形成群體,但是另一部分個體卻可能是孤獨生活的。例如:儘管大部分獅子以家族方式進行集群生活,但是另一些個體則是孤獨生活著。
分類
根據集群後群體持續時間的長短,可以把集群分為臨時性和永久性兩種類型。
①永久性集群
永久性集群存在於社會動物(蜜蜂、螞蟻、白蟻等)和高等動物(靈長目動物)。社會昆蟲由於分工專化的結果,同一物種群體的不同個體具有不同的形態。例如,在螞蟻社會當中,有大量的工蟻和兵蟻以及一隻蟻后,工蟻專門負責採集食物、養育後代和修建巢穴;兵蟻專門負責保衛工作,具有強大的口器;蟻后則成為專門產卵的生殖機器,具有膨大的生殖腺和特異的性行為,採食和保衛等機能則完全退化。
②臨時性集群
大多數的集群屬於臨時性集群,臨時性集群現象在自然界中更為普遍,如遷徙性集群、繁殖集群等季節性集群以及取食、棲息等組成的臨時性群體。
產生原因
生物產生集群的原因複雜多樣。主要包括以下方面:
①對棲息地的食物、光照、溫度、水份的共同需要。例如:潮濕的生境使一些蝸牛在一起聚集成群,一隻鹿的屍體作為食物和隱蔽地,招攬來許多食腐動物而形成群體;
②對晝夜天氣或季節氣候的共同反應。例如非洲草原上的角馬大遷徙;
③繁殖的結果,由於親代對某環境有共同的反應,將後代(卵或仔)產於同一環境,後代由此一起形成群體。例如大西洋鮭魚的洄游產卵後幼鮭魚的集群。家族式的集群也是由類似原因所引起的,但是家族當中的個體之間具有一定的親緣關係;
④被動運送的結果。例如強風、急流可以把一些蚊子、小魚運送到某一風速或流速較為緩慢的地方,形成群體;
⑤由於個體之間社會吸引力相互吸引的結果。集群生活的動物,尤其是永久性集群動物,通常具有一種強烈的集群欲望,這種欲望正是由於個體之間的相互吸引力所引起的。當一隻離群的鴿子,遇到一群素不相識的鴿子時,毫無疑問,這隻離群的鴿子將很快地加入到素不相識的鴿子群當中。有時候,由於強烈的聚群欲望,離群的個體在沒有其他同種生物可以聚群時,有些動物甚至加入到其他物種的群體,以滿足其聚群欲望,如離群的海鷗加入到海燕群中;
動物群體的形成可能是完全由環境因素所決定的,也可能是由社會吸引力所引起,根據這兩種不同的形成原因,動物群體可分為兩大類,前者稱為集會,後者稱為社會。
集群效應與生態學意義
動物界許多動物種類都是群體生活的,說明群體生活具有許多方面的生物學意義,群體優點的適應價值促進了動物社會結構的進化,目前已經知道許多種昆蟲和脊椎動物的集群能夠產生有利的作用。同一種動物在一起生活所產生的有利作用,成為集群效應。集群的生態學意義主要有以下幾個方面:
①集群有利於提高捕食效率;
②集群可以共同防禦敵害;
③集群有利於改變小生境;
④集群有利於某些動物種類提高學習效率;
⑤集群能夠促進繁殖。
領域性
若動物保衛一區域,不允許其他動物(通常是同種動物)進入,那么該區域或空間就稱為領域,而動物占有領域的行為則稱為領域性。領域行為是種內鬥爭的表現之一。
一些領域是暫時的,例如大部分鳥類都只是在繁殖期間才建立和保衛領域。一些領域則是永久的,如大型貓科食肉動物建立的領域。同種動物的資源需求相同,領域性由種內鬥爭引起,領土的所有者排斥其他相同物種個體的進入,這樣能減少競爭,並獲得更多的資源。此外,行為相似的物種間也有領域性行為,但這已經屬於群落範疇 。
社會等級
社會等級是指動物種群中各個動物的地位具有一定順序的等級現象。社會等級形成的基礎是支配行為,或稱支配—從屬關係。例如,家雞飼養者很熟悉雞群中的彼此啄擊現象,經過啄擊形成等級,穩定下來後,低級的一般表示妥協和順從,但有時也通過再次格鬥而改變順序等級。穩定的雞群往往生長快,產蛋也多,其原因是不穩定雞群中個體間經常的相互格鬥要消耗許多能量,這是社會等級制在進化選擇中保留下來的合理性的解釋。社會等級優越性還包括優勢個體在食物、棲所、配偶選擇中均有優先權,這樣保證了種內強者首先獲得交配和產後代的機會,所以從物種種群整體而言,有利於種族的保存和延續。
通訊
社會組織的形成,還需要有個體之間的相互傳遞信息為基礎。信息傳遞,或稱通訊是某一個體傳送信號,另一個體接受信號。並引起後者反應的過程。根據信號的性質和接受的感官,可以把通訊分為視覺的,化學的和聽覺的等。信息傳遞的目的很廣,如個體的識別,包括識別同種個體,同社群個體,同家族個體等,親代和幼仔之間的通訊,兩性之間求偶,個體間表示威嚇、順從和妥協,相互警報,標記領域等。從進化觀點而言,所選擇的應以傳遞方便、節省能量消耗,誤差小、信號傳送者風險小,對生存必須的信號。
種群與進化
概述
在現代生物進化理論中,種群是生物進化的基本單位,種內個體通過不定向變異與自然選擇不斷進化 。
基因庫
一個種群中全部個體的全部基因,就被稱為該種群的基因庫。在一個種群的基因庫中,某個基因占全部等位基因的比例,叫做基因頻率。
例如:在豌豆中,決定豌豆顏色的等位基因為:R(顯性,綠色)r(隱形,黃色)。假設從一個豌豆種群中隨機抽取200個個體,測得基因型為RR、Rr、rr的個體數分別為80、40、80,那么,可以計算得出:
R的基因數量=80×2+40=200(個)
r的基因數量=40+80×2=200(個)
R的基因頻率=200÷(200+200)=0.5
r的基因頻率=200÷(200+200)=0.5
註:此處僅是舉例說明,並非真實情況
哈代-溫伯格定律
哈代-溫伯格定律,也稱“遺傳平衡定律”,1908年,英國數學家戈弗雷·哈羅德·哈代( Godfrey Harold Hardy)最早發現並證明這一定律;1909年,德國醫生威廉·溫伯格( Wilhelm Weinberg)也獨立證明此定律,故得名。哈代-溫伯格定律主要用於描述群體中等位基因頻率以及基因型頻率之間的關係。內容為:
①一個無窮大的群體在理想情況下進行隨機交配,經過多代,仍可保持基因頻率與基因型頻率處於穩定的平衡狀態。
②在一對等位基因的情況下,基因p(顯性)與基因q(隱形)的基因頻率的關係為:
(p+q)^ 2=1
二項展開得:p^ 2+2pq+q^ 2=1
可見,式中“p^ 2”為顯性純合子的比例,2pq為雜合子的比例,“q^ 2”為隱形純合子的比例。
哈代-溫伯格定律在多倍體等更加複雜的情況下也可套用。
地理與生殖隔離
主條目:生殖隔離、地理隔離
 Kaibab松鼠(左)與Abert松鼠
Kaibab松鼠(左)與Abert松鼠不同種群的個體,在自然條件下無法相互交配或相互交配無法產生可育後代(如驢與馬雜交產生騾)的情況被稱作生殖隔離,生殖隔離可以區分不同物種或亞種,也就是說,生殖隔離是區分物種的標誌。
生殖隔離往往由地理隔離產生,同一種群,因地理因素(造山運動、大陸漂移等)被強行分開後,經過數億萬年的變異與自然選擇,就會形成不同的物種,比如美國科羅拉多大峽谷兩側的Kaibab松鼠與Abert松鼠,原先就是一個物種,後因為大峽谷河流的形成,它們的祖先被分為兩個隔離的種群,最後,形成了今天的樣子。
基因頻率
一個種群的全部個體所含有的全部基因,叫做這個種群的基因庫。種群中每個個體所含有的基因,只是種群基因庫的一個組成部分。某個基因在某個種群基因庫中所占的比例,叫做該基因的頻率。所有不同基因在種群基因庫中出現的比例,就組成了種群基因頻率。在自然界中,由於存在基因突變、基因重組和自然選擇等原因,種群基因頻率總是在不斷變化的。自然選擇實際上是選擇某些基因,淘汰另一些基因,所以自然選擇必然會引起種群基因頻率的定向改變,決定了生物進化的方向。
