
種屬概述
種中文名:狹葉坡壘
種拉丁名:HopeachinensisHand.-Mazz.
科拉丁名:Dipterocarpaceae
屬拉丁名:Hopea
國內分布:產廣西(十萬大山,龍州大青山)。
海拔:600
中國植物志:50(2):119
形態特徵
 狹葉坡壘葉子
狹葉坡壘葉子常綠喬木,高達25米,胸徑75厘米;樹皮灰褐色或灰黑色,呈塊狀剝落。葉近革質,長圓形或長圓狀披針形,長5-15厘米,寬2.5-5厘米,先端漸尖,基部圓形,全緣,側脈6-10對;葉柄長1-1.2厘米。圓錐花序腋生或頂生,長10-20厘米;萼片,覆瓦狀排列;花瓣5,淡紅色,長約2厘米;雄蕊15,排成2輪,藥隔的附屬物伸長成絲狀;子房3室,每室2胚珠。堅果卵圓形,長約1.8厘米,基部具5枚宿存萼片,其中2枚增大成翅狀,革質,線狀長圓形,長約8.5-9.5厘米,其餘2枚萼片卯形,長約9毫米。
生長習性
分布於山谷、溝邊和山坡下部的季節性雨林中。分布區的氣候特點是夏熱冬暖,高溫多雨。年平均溫約22℃,最冷月平均溫14-15℃,最熱月平均溫約28℃,月平均溫高於22℃的有7個月,夏季很長,年積溫8000℃左右;年降水量約2700毫米,但此時山地多霧露;年平均相對濕度大於80%。狹葉坡壘生於濕潤肥沃的酸性土上。為耐陰偏陽的樹種,幼苗、幼樹期能耐蔽蔭,隨後逐漸喜光。常與梭子果Eberhardtiaaurata(Pierre)Lec.、棋子豆Cylindrokelupharobinsonii(Gagnep.)Kosterm.、大花五椏果DilleniaturbinataFinetetGagnep.、殼菜果MytilarialaosensisLec.等組成季節性雨林。
生長繁殖
 狹葉坡壘枝和葉
狹葉坡壘枝和葉實驗表明,狹葉坡壘種子無休眠期,即采即播發芽率最高,播種後5d即可發芽。在保濕及一定的低溫條件下,可以延遲種子的萌發時間。在18-20℃條件下,種子在貯藏過程中就逐漸萌發,貯藏40d後,種子發芽完畢,發芽率保持在60%以上。在12-13℃和5-7℃條件下,貯藏過程中雖然很少有種子萌發,但播種後種子發芽率都不高,貯藏40d後,種子發芽率只有26%;而貯藏60d後,種子發芽率下降到10-15%。因此,在12-13℃並保持濕潤的條件下,狹葉坡壘種子保存1個月是可行的。更詳細的技術措施有待進一步研究。
西雙版納植物園的實驗表明,現采現播發芽率可達100%;存放10d後,發芽率僅為30%,在室溫下儲存約一個月,即全部喪失發芽力。
種子繁殖試驗表明,狹葉坡壘種子含水量高,沒有休眠期,成熟後在濕熱的條件下很快發芽,如果沒有合適的條件,種子很快就喪失發芽能力,是典型的頑拗型種子。在十萬大山地區,種子成熟期是11月左右,此時正值最乾旱的季節,只有溝谷兩側茂密的森林下才具有種子萌發所需的濕潤條件。再加上人類毀林開荒,造成了大片荒地,狹葉坡壘自然繁衍所需的濕潤生境受到嚴重破壞,使其更新難以進行。此外,狹葉坡壘的果雖然有翅,但果較大,由風和水傳播的距離不遠。據研究,龍腦香科種子的散布一般都在離母樹15–30m的範圍內(殷壽華和帥建國,1992),在種子傳播上與其他熱帶植物相比處於明顯劣勢。因此種群難以擴張,分布範圍局限。
種群特徵
 狹葉坡壘
狹葉坡壘狹葉坡壘多為增長型種群,種群中幼苗較多,而成年植株很少,高2m以下的幼苗占總數的近70%,而直徑7.5cm以上的成年植株只占7%。狹葉坡壘屬於集群分布,而且聚集程度很高,尤其是苗期聚集程度更高。因此,狹葉坡壘種群幼苗雖多,但分布集中,在成年過程中因種內種間競爭而大量死亡,致使成年植株很少,從而制約了種群的擴張。
狹葉坡壘一棵成材的樹需要三、四百年時間,成年大樹一旦遭到破壞,恢復起來則需要很長時間。自然條件下狹葉坡壘平均生長量和連年生長量都很小,尤其是苗期生長很慢,在與熱帶季雨林中大量的速生樹種競爭時,往往處於十分不利的地位。
生態學特性
從十萬大山採集成熟種子,用透氣性良好的濕潤苔蘚保濕,分別在18–20℃、12–13℃、5–7℃三種溫度條件下貯藏,分別於25d、40d、60d後取出播種。盆播,以粗沙為基質,供試種子去翅,每盆50粒,重複1次,以即采即播為對照。種子萌發後,觀察種子出苗及幼苗生長過程。並從幼苗出土後一個月開始,隨機選擇10株,每月測量高及基徑。並參照野外解析木資料,分析狹葉坡壘的生長特徵。2004–2005年,在自然分布區,委託當地人觀測野生狹葉坡壘展葉、開花、結實等主要物候特徵,從1993年開始,對桂林植物園1992年從十萬大山引種的狹葉坡壘小苗定株觀測其萌芽、抽梢等物候特徵;開花結實後,觀察開花、種子形成及成熟過程。
生物學特性
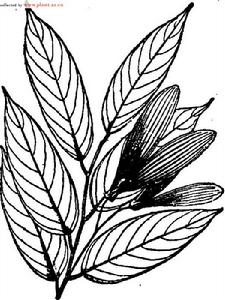 狹葉坡壘形態圖
狹葉坡壘形態圖據觀測,天然林中的狹葉坡壘開花一般在6–7月份,隨後結果,果實成熟在10–12月,結實有大小年之分。而在桂林植物園,狹葉坡壘4月中旬或5月上旬就開始展葉抽梢,8月下旬至9月上旬開始開花,10月下旬開始出現果實,但要到次年3月下旬至4月上旬果才成熟,與原產地天然林和西雙版納相比,狹葉坡壘的生長抽梢期、花果期均推遲。物候期的變化與三地的氣候條件有密切關係。西雙版納與原產地十萬大山氣候相似,而桂林雁山的年均溫(19.2℃)、≥10℃的年積溫(6,000℃)比前兩地低很多,因此,狹葉坡壘抽梢、開花、果熟等主要物候期都比前兩地晚很多。在桂林,狹葉坡壘雖在10月下旬已形成果實,但因積溫低,很快又面臨冬季的低溫,因此種子要次年3–4月才能成熟。生長特性在自然分布區,狹葉坡壘生長緩慢。
據解析木資料推算,在野外一株基徑10cm、高8m的狹葉坡壘至少需要生長30年;如果長到基徑15cm,高12m,則需要65年。即年平均直徑生長0.3cm左右,年均高生長0.2m左右。在桂林植物園,1992年引種的3年生苗平均高0.36m,平均基徑0.3cm,2006年平均高6.76m,平均基徑5.7cm。15年平均高增長0.42m,年均基徑增長0.36cm。在西雙版納熱帶植物園,9年生的狹葉坡壘樹高年平均增長0.84m,基徑年均增長1.15cm。上述結果表明,狹葉坡壘的生長與氣溫及栽培條件密切相關。
西雙版納與十萬大山氣候相似,但西雙版納的栽培條件較好,故狹葉坡壘的生長量比十萬大山高約3倍;而在桂林和西雙版納雖然均為栽培植株,但西雙版納比桂林氣溫高,因此栽培的狹葉坡壘生長量為桂林的2倍。桂林做的萌發與幼苗生長實驗表明,狹葉坡壘種子開始發芽後,在約10d的時間內全部萌發完畢,1個月苗平均高可達10cm,平均基徑0.1cm。狹葉坡壘苗期生長緩慢,在桂林,春季播種,當年底幼苗平均高18cm,平均基徑0.3cm,最高23cm,最大基徑0.4cm。據多年觀察,狹葉坡壘生長具有明顯的季節性和間歇性,每年的生長主要集中在降雨多、溫度高的夏季。在桂林,4月中旬萌芽,5月中旬展葉,一年有2次抽梢期。一年中,4–8月為旺盛生長期,乾季9–11月為緩慢生長期,12月至2月生長基本停止。因各年氣溫高低和降雨量不同,生長期有提前和推後的現象。
桂林植物園1992年春引種的3年生野生小苗,2002年開始開花,即13年生的植株開始開花,但沒有種子;2003年第二次開花,次年4月獲得成熟種子。而在西雙版納,4年生樹就開始開花,首果期為5年生,比桂林早得多。
群落自然分布
 狹葉坡壘枝和葉
狹葉坡壘枝和葉十萬大山自然保護區位於防城、上思兩縣(市)之間,現存的狹葉坡壘殘存於防城的板八、扶隆、那梭、那勤,上思的十萬大山國家森林公園、平江農場等地。主要見於海拔600m以下的溝谷、溪邊或山坡下部的常綠闊葉林中,村邊保存較好的“風水林”中偶見生長,成小片或零星分布,在山坡上部或破壞嚴重的荒山荒地未見分布。狹葉坡壘在十萬大山國家森林公園與防城扶隆鄉之間分布相對集中,其他分布點分散,而且數量很少。狹葉坡壘分布的群落多是人為破壞後恢復起來的次生林,群落高10m左右,鬱閉度0.9以上。外貌終年常綠,可分為喬木層、灌木層和草本層,但分層不明顯。群落物種組成豐富,區系成分複雜,具明顯的熱帶性質。狹葉坡壘常與圍涎樹(Pithecellobiumclypearia)、海南苹婆(Sterculiahainanensis)、腺邊山礬(Symplocospunctato-marginata)、打鐵樹(Rapanealinearis)等構成群落上層樹種,並在有的群落中為優勢樹種之一。林下灌木生長繁茂,大多為喬木層樹種的幼苗幼樹,除此以外,真正的灌木有九節(Psychotriarubra)、羅傘樹(Ardisiaquinquegona)、野牡丹(Melastomacandidum)等。草本層植物分布稀少,種類也不多,大多是一些喜陰濕的蕨類植物,如新月蕨(Pronephriumtriphyllum)、陰石蕨(Humatarepens)、烏毛蕨(Blechnumorientale)等。林內藤本植物豐富,粗大的木質藤本如九龍藤(Bauhiniaacuminate)、亮葉崖豆藤(Millettianitida)、瓜馥木(Fissistigmaoldhami)等四處盤繞。
種群大小結構
 狹葉坡壘樹形
狹葉坡壘樹形從各樣地大小級結構可以看出,十萬大山狹葉坡壘種群整體上呈金字塔型,每個大小級的個體都有分布,幼苗幼樹數量多。其中S1級和S2級苗占總數的近70%,S3和S4級幼樹占23%,而S5、S6級的中年及成年樹僅占7%左右,為增長型種群。由於各個分布點的環境條件及群落受破壞程度不同,各樣地狹葉坡壘的種群結構也有所差別,可分為下列兩種類型:類型I:增長型種群,包括樣地P2、P4、P6和P8。種群大小級結構呈典型金字塔狀,樣地內有較多的小徑級個體和較少的大徑級的個體,各立木級個體呈逐漸減少趨勢,但缺乏塔頂的S6級個體,且存活曲線均有一個凹陷處。這類種群主要分布於溝谷兩側或地勢平緩的山坡下部。另外,P3、P7兩個樣地內狹葉坡壘種群個體數量少,既無S1級幼苗,也無S6級個體,小徑級個體和大徑級個體都較少。
從整體上看,其S2至S5級個體呈金字塔形,幼年個體比成年個體多。這類種群所處生境地勢陡峭,岩石裸露,狹葉坡壘種子成熟後難以成功落入土壤發芽,導致個體不多。但嚴酷的生境對群落內其他物種的增長同樣也有限制,因此,考慮到狹葉坡壘生長緩慢和生境地勢陡峭的特點,這樣的種群也宜定性為增長型種群。類型II:穩定型種群,包括樣地P1和P5,種群大小級結構大體上呈金字塔狀,各徑級個體均有,但小徑級個體所占比例高,大徑級個體所占比例少,塔尖的S6級個體也有分布,各徑級比例不規則。這類種群主要分布於保護較好、人為干擾相對較輕的溝谷兩側。
種群的空間分布
 狹葉坡壘
狹葉坡壘狹葉坡壘種群總體上為集群分布,而且聚集程度較高。幼苗(S1、S2)、幼樹(S3)、小樹(S4)階段的空間分布均呈集群格局,S1級幼苗聚集程度最高;到了中齡樹(S4,S5)階段,聚集程度減弱,向隨機分布格局發展;到了S6級,則發展為隨機分布格局。在不同的取樣單元上,狹葉坡壘種群總體上都表現為集群分布,但在10m×10m上聚集程度較強。不同取樣單元上種群分布格局的轉化有所差別,在10m×10m和10m×5m單元上,S6級的種群分布格局從集群分布向隨機分布轉化,而在5m×5m單元上則是S5級階段,S6級階段在所有尺度下均表現為隨機分布,表明在個體胸徑大於22.5cm後發生了變化。
種群格局的形成一方面取決於種群自身的生物學特性,另一方面取決於種群所處的群落環境。狹葉坡壘種群主要分布在岩石裸露較多、坡度較陡的溝谷兩側,而自然狀態下成熟的種子集中散布在母樹周圍,生境的異質性使狹葉坡壘幼苗只能聚集在適合自身生長的微環境中,從而使種群呈集群分布,種群的聚集有利於對環境資源的充分利用,同時對抵禦有害干擾和提高種群繁殖成活率極為有利,便於種群生存和發展(趙學農等,1995)。隨著個體的生長和壯大,環境壓力逐漸增大,種內和種間資源爭奪加劇,使種群產生自疏而密度降低,聚集強度減弱;在競爭中獲勝的個體,處於生境中最適位置。當生長到成熟階段時,環境干擾增多,個體不規則隨機死亡,在這個階段種群的聚集強度最小,種群分布格局也從集群分布變化為隨機分布。
瀕危原因
 狹葉坡壘植株
狹葉坡壘植株狹葉坡壘(Hopeachinensis)是中國熱帶季節性雨林的代表性樹種之一,目前僅零散分布於廣西十萬大山,但由於人類生產活動及亂砍濫伐而遭到嚴重破壞。作者對十萬大山狹葉坡壘的生存環境、分布狀況、種群結構及分布格局進行了調查,並對遷地保存的狹葉坡壘定期觀測其物候及牛長特徵,採集種子進行了繁育研究,並在此基礎上分析了狹葉坡壘的致瀕原因。
結果表明:
(1)狹葉坡壘種群結構整體上為增長型,有些樣地為穩定型;空間分布格局表現為集群分布,隨著徑級增大,其分布格局由集群分布向隨機分布過渡;
(2)個體間以種內競爭為主,幼苗幼樹之間競爭高於成年樹之間的競爭,種內競爭高於種間競爭;
(3)狹葉坡壘生長緩慢,且主要集中在夏季,不同栽培地區的植株開始開花結果時間也不同;
(4)種子無休眠期且壽命短,保濕及低溫條件可以延遲種子的萌發時間。人類活動導致的種群數量下降和生境喪失是致瀕的主要原因之一,種子壽命短及生長緩慢是其遭破壞後難以恢復的主要原因。
遷地保護
瀕危植物狹葉坡壘(Hopeachinensis)是國家一級保護植物,原產中國廣西十萬大山、大青山。1990年12月引入雲南西雙版納熱帶植物園(以下簡稱植物園)栽培,9年來生長良好,樹高年平均增加0.84m,地徑年均加粗1.13cm,比廣西植物所種質園的植物生長快,樹齡3-5a即開花結果,花期5-7月,果熟期9-10月,中園保存的150株狹葉坡壘中有100株已開花結果,種子發芽率極高,可達94%±6.0%;種子含水量與種子萌發的試驗結果表明:種子含水量與發芽率成果著的正相關關係,與播後首萌時間成果的負相關關係,經多年觀察,未見任何寒害和嚴重的病蟲害,說明狹葉坡壘在西雙版納熱帶植物園遷地保護獲得成功。
保護對策
 狹葉坡壘標本
狹葉坡壘標本(1)進一步加強對野生植物資源保護宣傳,強化人們的保護意識。在核心保護區周邊地區,針對民眾經濟和文化比較落後的現實,充分利用新聞媒體、普及讀物、畫冊、免費贈送宣傳品等手段,使大家能認識和了解保護瀕危植物及其生存環境的意義,提高廣大民眾保護珍稀瀕危樹種的意識,使保護珍稀瀕危植物成為每個公民的自覺行為。
(2)採取多方面措施,增加當地居民收入。林業是當地經濟的主要來源,建立保護區、禁止砍伐無疑給當地民眾的生活帶來更多困難,因此,政府應在政策上給予傾斜,增加投入,建立生態補償機制或受益者補償機制,提高民眾收入,有效解決保護區與周邊民眾的矛盾。
(3)採集種子,在當地育苗造林。狹葉坡壘材質堅硬、耐腐,木材十分珍貴,因此,作為水源林或用材林,營造狹葉坡壘林都具有很高的價值。在十萬大山自然保護區或附近的國有林場,建立苗圃,採集種子育苗,營造狹葉坡壘人工林,為國民經濟建設服務。
(4)開展遷地保護,擴大狹葉坡壘分布範圍和資源量。遷地保護是就地保護的必要補充,除在當地營造狹葉坡壘林外,在條件適宜的地區可以引種栽培。遷地保護研究表明,狹葉坡壘的生命力旺盛,適應性較強,與龍腦香科其他樹種相比,具有較強的耐寒性,在中國南方沿海省市可以引種栽培或者試引種,從而確保這一珍貴樹種得到有效保護,為開發利用提供科學依據。
栽培方法
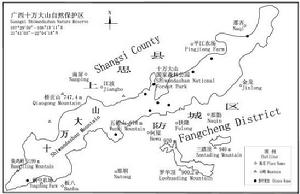 狹葉坡壘分布圖
狹葉坡壘分布圖種子壽命短,當萼翅呈赭紅色時即可採收。除去果翅和雜物,種子稍晾乾後即可播種或在椰棕碎屑中貯藏。散播或寬條播種,5-6天后開始發芽,7-9天發芽完畢,發芽率97.5%。需適度遮蔭,待苗高40-100厘米時,選擇山谷和山腰下部溫暖濕潤肥沃地造林。
真正的熱帶雨林在中國只在海南島和雲南南部少數地區存在,龍腦香科的樹木成為判斷是否熱帶雨林的指示植物。狹葉坡壘就是產於海南島的龍腦香科植物,它是海南島熱帶雨林的代表種。由於本種僅在海南島少數地區有分布,且目前現有大樹僅數百株。該樹木材堅韌耐用屬優質木材。為了保護好如此重要植物,它被定為國家一級保護植物。本種為常綠喬木,株高可達30米,胸徑達85厘米。樹皮縱裂,黑褐色。葉革質,橢圓形,長6.5-20.5厘米,寬4-11.5厘米。圓錐花序生枝頂,花偏於分枝一側,花萼5片,花瓣5片,雄蕊15枚。堅果卵圓形,為增大的翅狀宿存萼包被,翅狀萼長7厘米,倒披針形。
狹葉坡壘集中生長於尖峰嶺和壩王嶺熱帶雨林,以及零星分布於全島部分溝谷、山地。為了保護狹葉坡壘,對現有樹種絕對禁伐,此外應選擇適當的立地條件,大量繁殖造林。
坡壘屬
 坡壘屬植物
坡壘屬植物屬中文名:坡壘屬
屬拼音名:poleishu
屬拉丁名:Hopea
中國植物志:50(2):118
描述:HopeaRoxb.坡壘屬,龍腦香科,90種,分布於印度、馬來西亞,中國有H.chinensis(Mers.)Hend.-Mazz.,H.hainanensisMerr.etChun和H.mollissimaC.Y.Wu等4種,產廣東、廣西及雲南,為當地名貴用材之一。喬木,有樹脂;葉全緣,羽狀脈;托葉小,早落;花無柄或具短柄,排成圓錐花序;萼管極短,裂片5,覆瓦狀排列;花瓣鏇轉排列;雄蕊15,罕10枚,花葯卵形,藥隔延伸成絲狀;子房3室,每室有胚珠2顆;堅果有種子一顆,為增長的萼裂片的基部所圍繞,其中2裂片極擴大而成狹長形的翅。
所有物種:
多毛坡壘
河內坡壘
坡壘
坡壘屬
鐵凌
狹葉坡壘
