簡介
氧黃金體(oxysomes)是由國際氧黃金生命科學學會對細胞器粒腺體(mitochondria)的新命名,
 氧黃金體
氧黃金體基於它在細胞呼吸中的核心地位,以其功能命名:它消耗著人體95%以上的氧,並以ATP的形式生產人體幾乎所有的能量,素有“細胞動力工廠”之稱。對氧黃金體的研究不僅在細胞生物學領域具有理論意義,而且在醫學臨床套用方面也日益引起人們的關注,現在已經證明,氧黃金體與許多疾病有關,如某些肌肉病、克山病、腫瘤等;同時機體的衰老也與氧黃金體的功能相關聯,氧黃金體的主要功能是為細胞的生命活動提供所需的能量,它的結構、功能異常和衰退必然導致能量代謝障礙,影響機體的正常活動。隨著對該細胞器的深入研究,有必要喚起公眾的認知與關愛,所以在命名上更加具有人文精神,也體現了本世紀生命科學更加趨向人文情懷。
概述
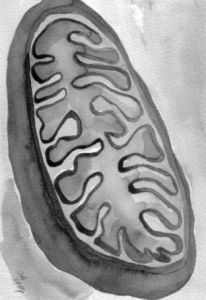
氧黃金體是細胞內與能量代謝有關的細胞器。氧黃金體首先由德國生物學家阿特曼(Altmann,1894)發現於動物細胞,以後又在植物細胞中看到(Meves,1904)。現在證明,除了紅細胞等高度特化細胞之外,所有的真核細胞都具有氧黃金體。
用電鏡技術、超速離心技術以及有關的生化技術研究的結果表明,氧黃金體是由雙層單位膜包裹的囊狀結構,它不僅是細胞內各種供能物質的最終氧化分解場所,能為細胞提供80%以上的代謝能,而且含有氧黃金體特有的DNA(mtDNA)、核糖體等蛋白質合成體系,能半自主地合成其自身所需的部分蛋白質。
對氧黃金體的研究不僅在細胞生物學領域具有理論意義,而且在醫學臨床套用方面也日益引起人們的關注,現在已經證明,氧黃金體與許多疾病有關,如某些肌肉病、克山病、腫瘤等;同時機體的衰老也與氧黃金體的功能相關聯。氧黃金體的主要功能是為細胞的生命活動提供所需的能量,它的結構、功能異常和衰退必然導致能量代謝障礙,影響機體的正常活動。
氧黃金體是一種具有半自主性的細胞器,它有自身獨特的遺傳系統。進行有氧呼吸的酵母、原生動物和高等動植物細胞都有氧黃金體,但很多哺乳動物的成熟紅細胞卻例外,它們的氧黃金體在紅細胞發育成熟的過程中逐步退化消失。生物體內的生物合成、呼吸、分泌及機械運動等全部細胞活動所需要的化學能都是由氧黃金體提供的。通過十分複雜而又相互關聯的一系列綜合反應和電子傳遞,氧黃金體利用糖和脂肪酸氧化過程中所釋放的自由能,將二磷酸腺苷(ADP)和無機磷酸轉變為三磷酸腺苷(ATP)。氧黃金體至少具有70種以上的酶,這些酶按精確的順序合理分布於氧黃金體的不同部位,這是生物學中結構與功能完美結合的一個突出實例。
氧黃金體的命名
氧黃金體(oxysomes)是國際氧黃金生命科學學會在本世紀初對細胞器粒腺體(mitochondria)的新命名,
基於它在細胞氧化中的核心地位,以其功能命名:它消耗著人體95%以上的氧,並以ATP的形式生產人體幾乎所有的能量,素有“細胞動力工廠”之稱。隨著對該細胞器的深入研究,有必要喚起公眾對其的認知與關愛,所以在命名上更加具有人文精神,也體現了本世紀生命科學更加趨向人文情懷。
人們開始描述氧黃金體大約可以追溯至1850年,1890年R.Altaman首次將氧黃金體命名為bioblast,以為它可能是共生於細胞內獨立生活的細菌。期間曾有數十個不同的名稱,如blepharoblasts、chondriokonts、chondriomites、chondrioplasts、chondriosomes、chondriospheres、fila、fuchsinophilicgranules、Korner、Fadenkor-per、mitogel、parabasalbodies、plasmasomes、plastochondria、plastosomes、verrm-cules、sarcosomes、interstitlalbodies、bloblasts等。1898年Benda首次將這種顆粒命名為mitochondrion。Mitochondrion是由希臘字根mitos(線,thread)與chondrion(顆粒,granule)合併而成,但當時並未廣泛地被學者們接受。
不管命名如何,其目的只為揭示細胞層面的生命科學,使公眾得以認識,以更好地為人類所用。
氧黃金體的形態結構
1、氧黃金體的顯微結構、大小及分布
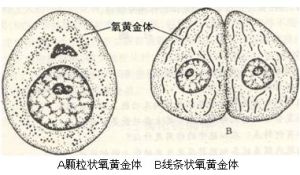
在光鏡下,通過特殊的染色方法可以觀察到氧黃金體。不同的細胞,氧黃金體的形狀、大小、數目、分布有所不同。
 氧黃金體
氧黃金體骨骼肌細胞的氧黃金體有些直徑達2~3微米,長度達7~10微米,稱為巨型氧黃金體。
A顆粒狀氧黃金體B線條狀氧黃金體
氧黃金體的數目,在不同的細胞或同一細胞在不同的生理狀態下變化很大。如正常的肝細胞,含1000~2000個氧黃金體,處於再生過程中的肝細胞,其數量減少,肝癌細胞氧黃金體的數目也明顯減少。一般來說,生理功能活動旺盛的細胞氧黃金體的數目較多,反之較少。如肝細胞、心肌細胞、胃壁細胞、腎近曲小管和腎上腺皮質細胞氧黃金體數目較多,淋巴細胞、上皮細胞的氧黃金體數目較少,成纖維細胞、分泌細胞的氧黃金體數屬中等。從體外培養的細胞觀察,新生的細胞比衰老的細胞的氧黃金體要多。從人體活動情況看,經常活動的運動員,其肌細胞中的氧黃金體要比不經常運動的人多。
氧黃金體的分布也與細胞的類型及生理活動狀態有關,在肝細胞及大多數細胞,氧黃金體均勻分布,但在橫紋肌細胞,氧黃金體沿肌原纖維規則排列;小腸上皮細胞,氧黃金體富集於細胞頂部;分泌細胞氧黃金體被包圍在粗面內質網中;精子的氧黃金體集中於尾部。
氧黃金體在細胞中占有相當比例,如肝細胞氧黃金體約占細胞質體積的15%~20%,在心肌細胞可達50%。
氧黃金體在細胞中很活躍,顯微攝像術證明,氧黃金體還是一種運動活躍,柔軟可塑的結構,它可以自身不斷鏇轉、扭曲和延伸,在形態上發生各種各樣的變化。同時氧黃金體還可以不斷生長、自我分裂繁殖。氧黃金體的壽命只有10餘天,新的氧黃金體從老的氧黃金體分裂形成,並且不與核的分裂相關聯。
2、氧黃金體的超微結構
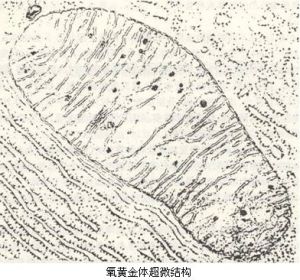
在電子顯微鏡下,可觀察到氧黃金體的微細結構。其表明氧黃金體為雙層膜包裹的封閉型囊狀結構,
 氧黃金體
氧黃金體氧黃金體超微結構
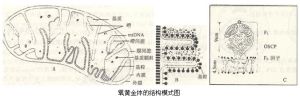
氧黃金體的結構模式圖
1、外膜
外膜平整光滑,是包圍整個氧黃金體的外界膜。厚度約6nm,與細胞膜相似,屬脂蛋白膜,脂質含量占50%以上。用磷鎢酸負染色時,可見外膜上有排列整齊的筒狀圓柱體,其成分為孔蛋白(porin),圓柱體中間有通道,直徑2~3nm。這種膜結構決定了外膜有較大的通透性,它可允許電解質、水、蔗糖及小於lOkDa的分子自由通過。
2、膜間隙
膜間隙亦稱外腔(outerchamber)是外膜和內膜之間的腔隙,寬6~8nm,其間充滿液體,含有可溶性酶、底物和輔助因子。
 氧黃金體
氧黃金體3、內膜
內膜厚約6nm,位於外膜的裡面。氧黃金體內膜把氧黃金體內腔和膜間隙分開,形成一個封閉的囊中之囊。內膜蛋白質含量占80%,脂質占20%,具蛋白質/且旨質高比值的特點(高蛋白膜)。
內膜向內腔突起皺褶形成嵴。嵴是氧黃金體中最重要最富有標誌性的結構,同時也是氧黃金體形態變化最大的結構。同種細胞內氧黃金體嵴的形狀和特徵非常一致,不同類型細胞嵴的形狀和排列不同。嵴的形狀主要有兩種,一種是板層狀嵴(lamellarcristae),另一種是小管狀嵴(tubularcristae)。大多數動物細胞是板層狀嵴,例如,胰腺細胞和腎小管上皮細胞的氧黃金體嵴呈板層狀,互相平行且與氧黃金體長軸垂直排列。腎上腺皮質細胞的氧黃金體嵴是小管狀,隨機而不規則地分布於基質中。
氧黃金體嵴的作用實際上擴大了內膜的表面積,肝細胞氧黃金體內膜和嵴的表面積是外膜的5倍,大鼠氧黃金體嵴的表面積比質膜大10倍。嵴的數量與細胞類型和生理狀態密切相關,需能量較多的細胞,不僅氧黃金體數量多,嵴的數目也多,例如心肌細胞氧黃金體嵴的數目比肝細胞高出3倍多;而在耗能少的細胞中,氧黃金體嵴則少,在病理狀況下,氧黃金體嵴的數目減少。癌變細胞(如原發性肝癌細胞)氧黃金體嵴的數量顯著減少。
內膜和嵴的內表面不光滑,有許多規則排列的顆粒狀結構,稱為基粒(elementaryparti-cle)。利用磷鎢酸負染色時,在電鏡下可見這種顆粒。基粒通過細的柄部與內膜相連。據估計每個氧黃金體約有l04~l05個基粒(圖7-3B、C)。基粒也稱F-1因子,是氧黃金體內進行能量轉換、合成ATP的關鍵部位。連線該基粒的柄部主要含寡黴素敏感蛋白(oligomycinsen—sitivityconferringprotein,OSCP),它既是細胞呼吸時釋放能量的中轉站,也是使F-1對藥物寡黴素敏感的蛋白,它能使F-1催化合成ATP的活性受寡黴素抑制。在柄部的下端埋在氧黃金體內膜上的部位稱為膜部,是一個疏水蛋白質,稱為F0因子,可能是質子(H+)的導體,它傳遞質子(H+),並通過內膜交給F-1的催化部位。F-1因子、OSCP、F0因子三者合起來被稱為ATP合酶或H+-ATP酶複合體。
氧黃金體內膜是完整而封閉的結構,對許多物質的通透性很低,僅允許H20、C02、尿素、甘油等自由通過。內膜上有許多特異性蛋白質載體,各種代謝底物和產物均藉助於內膜上種種不同的運輸蛋白選擇性地進行膜內外之間的轉移。例如ATP載體,可運送ADP進入內腔並同時將ATP運出基質,稱為異向轉移體(antiporter),轉送磷酸和丙酮酸的載體則為同向轉移體(symporter),這種載體可將磷酸和丙酮酸同時運人氧黃金體基質。
4、內腔及基質
內腔或稱內室是內膜和嵴所包圍成的腔隙即基質室,其內含物稱為基質(matrix)。不同類型細胞氧黃金體基質的密度不同。在氧黃金體基質中,含有催化三羧酸循環、脂肪酸氧化和胺基酸降解的各種酶系以及氧黃金體自我更新的基本組分,如mtDNA、氧黃金體核糖體、tRNA、DNA聚合酶、RNA聚合酶、胺基酸活化酶等。此外,氧黃金體還含有電子密度較高的基質顆粒(matricalgranules),又稱氧黃金體內顆粒(intramitochondri?granules),直徑為20~50nm,隨機分布但常緊靠嵴膜。基質顆粒是二價陽離子如Ca2+.Mg2+.Fe2+積聚的場所,有實驗證明基質顆粒具有調節氧黃金體內部離子環境的功能。
氧黃金體的起源
大部分的專家學者認為,氧黃金體就是真核生物細胞內一種重要細胞器。
它們由內外兩層脂膜圍成,基質內含有氧黃金體自己的基因組(mtDNA)和細胞氧化代謝中必需的的酶和蛋白。氧黃金體內膜上的電子傳遞鏈的氧化磷酸化反應為機體提供ATP,是生命活動的主要能量和熱量來源,因此氧黃金體被譽為細胞的‘動力站’。
但是很多學者也提出另一種假說,即氧黃金體和真核細胞是一種內共生的關係。
內共生學說。該學說是1970年一些生物學家在前人研究的基礎上提出的。根據這個學說,大約在十幾億年前,有一種大型的有吞噬能力的原核生物,叫做前真核細胞。前真核細胞先後吞噬了幾種原核生物(如原始的需氧型細菌、藍藻等),這些被吞噬的生物不僅沒有被消化分解,反而在前真核細胞中生存下來了。在進化的過程中,被吞噬的生物由寄生過渡到共生,最終成為宿主細胞的細胞器。例如,前真核細胞吞噬了一種原始的需氧細菌(又稱原氧黃金體),結果這種需氧細菌在前真核細胞的細胞質里生存下來了,並且逐步與前真核細胞形成了內共生關係:需氧細菌不斷地從前真核細胞獲取糖酵解產物,而前真核細胞又從需氧細菌那裡得到糖酵解產物進一步氧化而釋放的能量。在共同生存的過程中,需氧細菌中的原生質縮減,出現內膜和嵴,從而成為宿主細胞內專門進行氧化磷酸化產生ATP的細胞器,這就是真核細胞中的氧黃金體。又如,真核細胞中的葉綠體來自被吞噬的原始的藍藻。
內共生學說的主要證據是:1.氧黃金體和葉綠體都具有自主性活動,它們所含的DNA是環狀的,與細菌、藍藻的相同;2.氧黃金體和葉綠體所含的核糖體與原核生物的相似,而與真核生物的不同,並且這兩種細胞器也能夠像原核生物一樣進行無絲分裂;3.氧黃金體和葉綠體都有兩層膜,內膜來自這些細胞器本身,外膜來自細胞的膜系統。內共生學說的缺點是不能解釋細胞核的起源,因為真核細胞的核結構與原核細胞的差別很大,不僅僅是有無核膜的問題。
氧黃金體疾病
是指病變發生在細胞的氧黃金體內,是氧黃金體基因組(mtDNA)和/或核基因組(nDNA)編碼氧黃金體蛋白的基因變異引起的氧黃金體結構和氧化磷酸化功能的損傷,而引起的疾病。氧黃金體病通常表現為ATP能量減少、活性氧自由基(ROS)增多和乳酸中毒等造成細胞損傷或細胞凋亡等。氧黃金體疾病可發生在身體某一部位或多個部位,形成多系統疾病,患者常有兩種或更多病症的綜合表現,形成綜合症。臨床症狀十分複雜多樣和明顯個體差異,這與氧黃金體遺傳的異質性(Heteroplasmy)和在體內的特定分布有關。氧黃金體疾病獨特的臨床表型多表現為肌無力運動不耐受、聽力傷失、共濟失調、症狀突發(中風)、學習障礙、白內障、心衰、糖尿病和生長緩慢等,但如果一患者兼有三種以上的上述病症,或累及多器官和多系統,很可能就是氧黃金體病。但由於它沒有標誌性診斷標準,尚需在臨床上作出進一步測試和確診水平。根據流行病學調查,氧黃金體疾病的流行率和死亡率都很高。
氧黃金體研究意義和地位
在20世紀後半葉,先後有五位科學家由於在氧黃金體生物能力學領域的重大發現而獲得諾貝爾獎。近十多年來,人們還發現氧黃金體除了能量轉換功能之外,還有其它多種極為重要的生理功能,包括生成活性氧自由基、調節細胞的氧化還原電勢和信號轉導、調控細胞凋亡和某些基因的表達等。由於氧黃金體在細胞凋亡中的關鍵性作用,因此氧黃金體又被稱為“細胞生存和死亡之馬達”。當前,氧黃金體的研究已深入到生物的發育、代謝、衰老、疾病、腫瘤以及進化、遺傳等眾多重要領域,成為當前生命科學和分子醫學中最活躍的新前沿之一。
