發現過程
1879年,W. Flemming提出了染色質(chromatin)這一術語,用以描述細胞核中能被鹼性染料強烈著色的物質。
1888年,Waldeyer正式提出染色體的命名。
經過一個多世紀的研究,人們認識到,染色質和染色體是在細胞周期不同階段可以相互轉變的形態結構。
成分
通過分離胸腺、肝或其他組織細胞的核,用去垢劑處理後再離心收集染色質進行生化分析,確定染色質的主要成分是DNA和組蛋白,還有非組蛋白及少量RNA。大鼠肝細胞染色質常被當作染色質成分分析模型,其中組蛋白與DNA含量之比近於1:1,非組蛋白與DNA之比是0.6:1,RNA與DNA之比為0.1:1。DNA與組蛋白是染色質的穩定成分,非組蛋白與RNA的含量則隨細胞生理狀態不同而變化。
染色質DNA
基因組
 染色質
染色質凡是具有細胞形態的生物其遺傳物質都是DNA,只有少數病毒的遺傳物質是RNA。在真核細胞中,每條未複製的染色體包含一條縱向貫穿的DNA分子。狹義而言,某一生物的細胞中儲存於單倍染色體組中的總遺傳信息,組成該生物的基因組。真核生物基因組DNA的含量比原核生物高得多。
突變分析結果表明,並非所有基因都是細胞生存的必需基因,如酵母基因組有40%的基因屬於非必需基因,果蠅基因組只有5000個必需基因,最小最簡單的細胞支原體,有迄今發現的能獨立生活的有機體的最小基因組(482個基因),其中只有256個必需基因。
類型
以人類基因組為例,生物基因組DNA可以分為以下幾類。
1、蛋白編碼序列。以三聯體密碼方式進行編碼。編碼DNA在基因組中所占比例隨生物而異,在人類細胞基因組中,這一比例只有1.5%左右。這類編碼序列主要是非重複的單一DNA序列,一般在基因組中只有一個拷貝(單一基因),然而,也有可能有兩個或幾個拷貝甚至多達上千個拷貝的情況,這些都來自於從基因家族裡派生出來的重複基因或多基因。
2、編碼rRNA、tRNA、snRNA和組蛋白的串聯重複序列。它們在基因組中一般有20~300個拷貝,人類基因組中約含有0.3%這樣的DNA。
3、含有重複序列的DNA。這類DNA在基因組中占有很大一部分。它們又可以分為兩個亞類:簡單序列DNA和散在重複序列。DNA轉座子、LTR反轉座子、非LTR反轉座子和假基因都屬於散在重複序列。非LTR反轉座子包括短散在元件和長散在元件。典型SINE其長度少於500bp,如人和靈長類基因組中大量分散存在的Alu家族,人基因組中有50萬~70萬份Alu拷貝,相當於平均每隔4kb就有一個Alu序列;典型LINE其長度在6~8kb之間,如人基因組中L1家族,有100 000個L1拷貝。
4、未分類的間隔DNA。
5、高度重複DNA序列:
①衛星DNA,重複單位長5~100bp,不同物種重複單位鹼基組成不同,一個物種也可能含有不同的衛星DNA序列。
②小衛星DNA,重複單位長12~100bp,重複3 000次之多,又稱數量可變的串聯重複序列,每個小衛星區重複序列的拷貝數是高度不變的,因此早前常用於DNA指紋技術作個體鑑定。研究發現小衛星序列的改變可以影響鄰近基因的表達,基因的異常表達會導致一系列不良後效應。
③微衛星DNA,重複單位序列最短,只有1~5bp,串聯成簇長度50~100bp。
二級結構
生物的遺傳信息儲存在DNA的核苷酸序列中,生物界物種的多樣性也寓於DNA分子4種核苷酸千變萬化的排列之中。DNA分子不僅一級結構具有多樣性,而且二級結構也具有多態性。所謂二級結構是指兩條多核苷酸鏈反向平行盤繞所生成的雙螺鏇結構。DNA二級結構構型分3種:
①B型DNA(右手雙螺鏇DNA),是“經典”的Watson-Crick結構,二級結構相對穩定,水溶液和細胞內天然DNA大多為B型DNA;
②A型DNA(右手雙螺鏇DNA),是一般B型DNA的重要變構形式,其分子形狀與RNA的雙鏈區和DNA/RNA雜交分子很相近;
③Z型DNA(左手雙螺鏇DNA),也是B型DNA的變構形式。
3種構型DNA中,特別是大溝的特徵在遺傳信息表達過程中起關鍵作用,基因表達調控蛋白都是通過其分子上特定的胺基酸側鏈與溝中鹼基對兩側潛在的氫原子供體(═NH)或受體(O和N)形成氫鍵而識別DNA遺傳信息的。由於大溝和小溝中這些氫原子供體和受體各異以及排列不同,所以大溝攜帶的信息要比小溝多。此外,溝的寬窄及深淺也直接影響鹼基對的暴露程度,從而影響調控蛋白對DNA信息的識別。B型DNA是活性最高的DNA構型,變構後的A型DNA仍有較高活性,變構後的Z型DNA活性明顯降低。
此外,DNA雙螺鏇能進一步扭曲盤繞形成特定的高級結構,正、負超螺鏇是DNA高級結構的主要形式。DNA二級結構的變化與高級結構的變化是相互關聯的,這種變化在DNA複製、修復、重組與轉錄中具有重要的生物學意義。
染色質蛋白
與染色質DNA結合的蛋白負責DNA分子遺傳信息的組織、複製和閱讀。這些DNA結合蛋白包括兩類:一類是組蛋白,與DNA結合但沒有序列特異性;另一類是非組蛋白,與特定DNA序列或組蛋白相結合。
組蛋白
 染色質
染色質組蛋白是構成真核生物染色體的基本結構蛋白,富含帶正電荷的Arg和Lys等鹼性胺基酸,等電點一般在pH10.0以上,屬鹼性蛋白質,可以和酸性的DNA緊密結合,而且一般不要求特殊的核苷酸序列。
用聚丙烯醯胺凝膠電泳可以區分5種不同的組蛋白:H1、H2A、H2B、H3和H4。幾乎所有真核細胞都含有這5種組蛋白,而且含量豐富,每個細胞每種類型的組蛋白約6×10個分子。5種組蛋白在功能上分為兩組:
①核小體組蛋白。包括H2A、H2B、H3和H4。這4種組蛋白有相互作用形成複合體的趨勢,它們通過C端的疏水胺基酸互相結合,而N端帶正電荷的胺基酸則向四面伸出以便與DNA分子結合,從而幫助DNA捲曲形成核小體的穩定結構。這4種組蛋白沒有種屬及組織特異性,在進化上十分保守,特別是H3和H4是所有已知蛋白質中最為保守的。從這種保守性可以看出,H3和H4的功能幾乎涉及它們所有的胺基酸,任何位置上胺基酸殘基的突變可能對細胞都將是有害的。
②H1組蛋白。其分子較大。球形中心在進化上保守,而N端和C端兩個“臂”的胺基酸變異較大,所以H1在進化上不如核小體組蛋白那么保守。在構成核小體時H1起連線作用,它賦予染色質以極性。H1有一定的種屬及組織特異性。在哺乳類細胞中,組蛋白H1約有6種密切相關的亞型,胺基酸順序稍有不同。在成熟的魚類和鳥類的紅細胞中,H1 為H5取代。有的生物如酵母缺少H1,結果酵母細胞差不多所有染色質都表現為活化狀態。
非組蛋白
非組蛋白主要是指與特異DNA序列相結合的蛋白質,所以又稱序列特異性DNA結合蛋白(sequence specific DNA binding protein)。利用凝膠延滯實驗(gel retardation assay),可以在細胞抽提物中進行檢測。首先製備一段帶有放射性標記的已知特異序列的DNA,將要檢測的細胞抽提物與標記DNA混合,進行凝膠電泳。未結合蛋白的自由DNA在凝膠上遷移最快,而與蛋白質結合的DNA遷移慢,一般結合的蛋白質分子越大,DNA分子的延滯現象越明顯,然後通過放射自顯影分析,即可發現一系列DNA帶譜,每條帶分別代表不同的DNA-蛋白質複合物。每條帶相對應的結合蛋白隨後再通過細胞抽提物組分分離方法被進一步分開。
特性
①酸鹼性:組蛋白是鹼性的,而非組蛋白則大多是酸性的。
②多樣性:非組蛋白占染色質蛋白的60%~70%,不同組織細胞中其種類和數量都不相同,代謝周轉快。包括多種參與核酸代謝與修飾的酶類如DNA聚合酶和RNA聚合酶、HGM蛋白(high mobility group protein)、染色體支架蛋白、肌動蛋白和基因表達蛋白等。
③特異性:能識別特異的DNA序列,識別信息來源於DNA核苷酸序列本身,識別位點存在於DNA雙螺鏇的大溝部分,識別與結合靠氫鍵和離子鍵。在不同的基因組之間,這些非組蛋白所識別的DNA序列在進化上是保守的。這類序列特異性DNA結合蛋白具有一個共同特徵,即形成與DNA結合的螺鏇區並具有蛋白二聚化的能力。
④功能多樣性:雖然與DNA特異序列結合的蛋白質在每一個真核細胞中只有10 000個分子左右,約占細胞總蛋白的1/50 000,但具有多方面的重要功能,包括基因表達的調控和染色質高級結構的形成。如幫助DNA分子摺疊,以形成不同的結構域;協助啟動DNA複製,控制基因轉錄,調節基因表達等。
結構模式
雖然非組蛋白種類眾多,但是根據它們與DNA結合的結構域不同,可分為不同的家族。
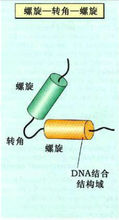
①α螺鏇-轉角-α螺鏇模式(helix - turn - helix motif)
 染色質
染色質這是最早在原核基因的激活蛋白和阻抑物中發現的。迄今已經在百種以上原核細胞和真核生物中發現這種最簡單、最普遍的DNA結合蛋白的結構模式。這種蛋白與DNA結合時,形成對稱的同型二聚體(symmetric homodimer)結構模式。構成同型二聚體的每個單體由20個胺基酸的小肽組成α螺鏇-轉角-α螺鏇結構,兩個α螺鏇相互連線構成β轉角,其中羧基端的α螺鏇為識別螺鏇(recognition helix),負責識別DNA大溝的特異鹼基信息,另一個α螺鏇沒有鹼基特異性,與DNA磷酸戊糖鏈骨架接觸。這種蛋白在與DNA特異結合時,以二聚體形式發揮作用,結合靠蛋白質的胺基酸側鏈與特異鹼基對之間形成氫鍵。
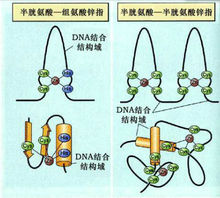
②鋅指模式(zinc finger motif)
 染色質
染色質負責 5S RNA、tRNA 和部分 snRNA 基因轉錄的RNA聚合酶Ⅲ所必須的轉錄因子。TFⅢ A 是首先被發現的鋅指蛋白,由344個胺基酸組成。TFⅢ A 含有9個有規律的鋅指重複單位,每個單位30個胺基酸殘基,其中一對半胱氨酸和一對組氨酸與Zn形成配位鍵。每個鋅指單位是一個DNA結合結構域(DNA-binding domain),每個鋅指的 C 末端形成α螺鏇負責與DNA結合。這類Cys/His鋅指單位的共有序列(consensus sequence)是:Cys - X - Cys - X - Leu - X - His - X - His。哺乳類轉錄因子 SP1 也有類似的鋅指結構,由三個鋅指單位組成。另一類鋅指蛋白含兩對半胱氨酸,而不含組氨酸,如哺乳類細胞的甾體類激素受體蛋白。這類Cys/Cys鋅指單位的結合Zn的共有序列是:Cys - X - Cys - X- Cys - X- Cys。不同的鋅指識別不同的鹼基序列,因為不同鋅指的胺基酸組成不一樣。
③亮氨酸拉鏈模式(leucine zipper motif,ZIP)
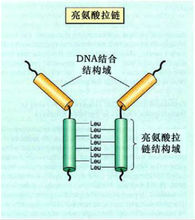 染色質
染色質在構建轉錄複合物過程中,普遍涉及蛋白與蛋白之間的相互作用,形成二聚體是識別特異DNA序列蛋白的相互作用的共同原則,亮氨酸拉鏈就是富含Leu殘基的一段胺基酸序列所組成的二聚化結構。這類序列特異性DNA結合蛋白家族,包括酵母的轉錄激活因子(GCN4)、癌蛋白Jun、Fos、Myc以及增強子結合蛋白(enhancer binding protein,C/EBP)等。所有這些蛋白的肽鏈羧基端約35個胺基酸殘基有形成α螺鏇的特點,每兩圈(7個胺基酸殘基)有一個亮氨酸殘基。這樣,在α螺鏇一側的Leu排成一排,兩個蛋白質分子的α - 螺鏇之間靠Leu殘基之間的疏水作用力形成一條拉鏈狀結構。這類蛋白與DNA的特異結合都是以二聚體形式起作用的,但與DNA結合的結構域是拉鏈區相鄰的肽鏈 N 端帶正電荷的鹼性胺基酸區。
④螺鏇-環-螺鏇結構模式(helix - loop - helix motif,HLH)
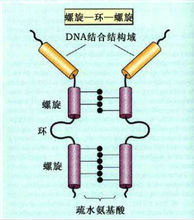 染色質
染色質HLH這一結構模式廣泛存在於動、植物DNA結合蛋白中。HLH由40~50個胺基酸組成兩個兩性α螺鏇,兩個α螺鏇中間被一個或幾個β轉角組成的環區所分開。每個α螺鏇由15~16個胺基酸殘基組成,並含有幾個保守的胺基酸殘基。具有疏水面和親水面的兩性α螺鏇有助於二聚體的形成。α螺鏇鄰近的肽鏈 N 端也有帶正電荷的鹼性胺基酸區與靶DNA大溝結合。具有螺鏇-環-螺鏇結構的蛋白家族成員之間形成同源或異源二聚體是這類蛋白與DNA結合的必要條件,缺失α螺鏇的二聚體不能牢固結合DNA。
⑤HMG框結構模式(HMG-box motif)
在發現一組豐富的高速泳動族蛋白(high mobility group protein)以後,首先命名HMG框結構模式。該結構由3個α螺鏇組成 boomerang-shaped 結構模式,具有彎曲DNA的能力。因此,具有HMG框結構的轉錄因子又稱為“構件因子(architectural factor)”,它們通過彎曲DNA、促進與鄰近位點相結合的其他轉錄因子的相互作用而激活轉錄。SRY是一種HMG蛋白,在人類男性性別分化中具有關鍵作用,HMG蛋白由Y染色體上一個基因編碼,在誘導睪丸分化途徑中一些相關基因的轉錄活性被HMG蛋白所激活。
結構
結構單位
20世紀70年代以前,人們關於染色質結構的傳統看法認為,染色質是組蛋白包裹在DNA外面形成的纖維狀結構。直到1974年Kornberg等人根據染色質的酶切和電鏡觀察,發現核小體是染色質組裝的基本結構單位,提出染色質結構的“串珠”模型,從而更新了人們關於染色質結構的傳統觀念。
實驗依據
 染色質
染色質1、用溫和的方法裂解細胞核,將染色質鋪展在電鏡銅網上,通過電鏡觀察,未經處理的染色質自然結構為30nm的纖絲,經鹽溶液處理後解聚的染色質呈現一系列核小體彼此連線的串珠狀結構,串珠的直徑為10nm。
2、用非特異性微球菌核酸酶消化染色質時,經過蔗糖梯度離心及瓊脂糖凝膠電泳分析,發現絕大多數DNA被降解成大約 200 bp的片段;如果部分酶解,則得到的片段是以 200 bp為單位的單體、二體、三體等。蔗糖梯度離心得到的不同組分,在波長 260 nm的吸收峰的大小和電鏡下所見到的單體、二體和三體的核小體組成完全一致。如果用同樣方法處理裸露的DNA,則產生隨機大小的片段群體。從而提示染色體DNA除某些周期性位點之外均受到某種結構的保護,避免酶的接近。
3、套用X射線衍射、中子散射和電鏡三維重建技術,研究染色質結晶顆粒,發現核小體顆粒是直徑為 11 nm、高 6.0 nm的扁圓柱體,具有二分對稱性。核心組蛋白的構成是先形成(H3)-(H4)四聚體,然後再與兩個H2A-H2B異二聚體結合形成八聚體。
4、SV40微小染色體分析。用SV40病毒感染細胞,病毒DNA進入細胞後,與宿主的組蛋白結合,形成串珠狀微小染色體,電鏡觀察SV40 DNA為環狀,周長1 500 nm,約 5.0 kb。若 200 bp相當於一個核小體,則可形成25個核小體,實際觀察到23個,與推斷基本一致。如用0.25mol/L鹽酸將SV40溶解,可在電鏡下直接看到組蛋白的聚合體,若除去組蛋白,則完全伸展的DNA長度恰好為 5.0 kb。
結構要點
1、每個核小體單位包括 200 bp左右的DNA超螺鏇和一個組蛋白八聚體以及一個分子的組蛋白H1。
2、組蛋白八聚體構成核小體的盤狀核心顆粒,相對分子質量100 000,由4個異二聚體組成,包括兩個H2A-H2B和兩個H3-H4。
3、146 bp的DNA分子超螺鏇盤鏇組蛋白八聚體1.75圈。組蛋白H1在核心顆粒外結合額外 20 bp DNA,鎖住核小體DNA的進出端,起穩定核小體的作用。
4、兩個相鄰核小體之間以連線DNA相連,典型長度 60 bp,不同物種變化值為 0~80 bp不等。
5、組蛋白與DNA之間的相互作用主要是結構性的,基本不依賴於核苷酸的特異序列。正常情況下不與組蛋白結合的DNA,當與從動、植物中分離鈍化的組蛋白共同孵育時,可以體外組裝成核小體亞單位。實驗表明,核小體具有自組裝的性質。
6、核小體沿DNA的定位受不同因素的影響。如非組蛋白與DNA特異性位點的結合,可影響鄰近核小體的相位;DNA盤繞組蛋白核心的彎曲也是核小體相位的影響因素,因為富含AT的DNA片段優先存在於DNA雙螺鏇的小溝,面向組蛋白八聚體,而富含GC的DNA片段優先存在於DNA雙螺鏇的大溝,面向組蛋白八聚體,結果核小體傾向於形成富含AT和富含GC的理想分布,從而通過核小體相位改變影響基因表達。
前期組裝
整個過程如下:
①最開始是H3·H4四聚體的結合,由CAF-1介導與新合成的裸露的DNA結合。
②然後是兩個H2A·H2B二聚體由NAP-1和NAP-2介導加入。為了形成一個核心顆粒,新合成的組蛋白被特異地修飾。組蛋白H4的Lys5和Lys12兩個位點典型地被乙醯化。
③核小體最後的成熟需要ATP來創建一個規則的間距以及組蛋白的去乙醯化。ISWI和SWI/SNF家族的蛋白參與此過程的調節。連線組蛋白(H1)的結合伴隨著核小體的摺疊。
④6個核小體組成一個螺鏇或由其他的組裝方式形成一個螺線管結構。
⑤進一步的摺疊事件將使染色質在細胞核中最終形成確定的結構。
 染色質
染色質這樣一個高度壓縮的結構極大地阻礙了像轉錄這樣的細胞核活動的進行。為了解決這個問題,有兩個家族的染色質修飾酶在染色質上作用,使染色質更接近於轉錄機器。第一個家族是通過在組蛋白尾部的共價修飾而發揮作用,這些修飾包括組蛋白的磷酸化、乙醯化和泛素化等,它們會影響以後與DNA或組蛋白相互作用因子的作用。第二個家族成員的主要特點是它們能夠利用ATP水解時釋放的能量來破壞核小體中的組蛋白-DNA接觸。
在真核生物細胞周期的S期,染色體的完全複製不僅需要基因組DNA的複製,也需要把複製好的DNA組裝成染色質。普遍認為,在複製叉的移動期間,染色質短暫地解組裝,然後在兩條複製好的子代DNA鏈上重新進行組裝。新複製的DNA主要通過以下兩種途徑組裝成染色質:第一,在複製叉的移動期間,父代的核小體核心顆粒與DNA分離,到該段DNA複製完成,父代的核小體核心顆粒直接轉移到兩條子鏈DNA的一條上;第二,染色質組裝因子利用剛剛合成的、乙醯化的組蛋白介導核小體在複製DNA上組裝。
染色質組裝的前期過程,即從裸露DNA組裝成直徑30納米的螺線管已有直接的實驗證據,並被絕大多數科學家認可。然而,染色質如何進一步組裝成更高級結構,直至最終成染色體的過程尚不是非常清楚,主要有兩種模型。
組裝模型
人的每個體細胞所含DNA約6×109bp分布在46條染色體中,總長達2米,平均每條染色體DNA分子長約5厘米,而細胞核直徑只有5~8微米,這就意味著從染色質DNA組裝成染色體要壓縮近萬倍,相當於一個網球內包含有2千米長的細線。
多級螺鏇模型
由DNA與組蛋白組裝成核小體,在組蛋白H1的介導下核小體彼此連線形成直徑約10納米的核小體串珠結構,這是染色質組裝的一級結構。不過在細胞中,染色質很少以這種伸展的串珠狀形式存在。當細胞核經溫和處理後,在電鏡下往往會看到直徑為30納米的染色質纖維。在有組蛋白H1存在的情況下,由直徑10納米的核小體串珠結構螺鏇盤繞,每圈6個核小體,形成外徑25~30納米,螺距12納米的螺線管。組蛋白H1對螺線管的穩定起著重要作用。螺線管是染色質組裝的二級結構。
Bak等(1977)從胎兒離體培養的分裂細胞中分離出染色體,經溫和處理後,在電鏡下看到直徑0.4微米,長11~60微米的染色線,成為單位線。在電鏡下觀察,判明單位線是由螺線管進一步螺鏇化形成直徑為0.4微米的圓筒狀結構,稱為超螺線管,這是染色質組裝的三級結構。這種超螺線管進一步螺鏇摺疊,形成長2~10微米的染色單體,即染色質組裝的四級結構。經過四級螺鏇組裝形成的染色體結構,共壓縮了8 400倍。
骨架-放射環結構模型
Laemmli等人用2mol/L的NaCl或硫酸葡聚糖加肝素處理HeLa細胞中期染色體,除去組蛋白和大部分非組蛋白後,在電鏡下可觀察到由非組蛋白構成的染色體骨架和由骨架伸出的無數的DNA側環。此外,實驗觀察發現,不論是原核細胞的染色體還是兩棲類卵母細胞的燈刷染色體或昆蟲的多線染色體,幾乎都含有一系列的袢環結構域,從而提示袢環結構可能是染色體高級結構的普遍特徵。
該模型認為,30納米的染色線摺疊成環,沿染色體縱軸,由中央向四周伸出,構成放射環,即染色體的骨架-放射環結構模型。首先是直徑2納米的雙螺鏇DNA與組蛋白八聚體構建成連續重複的核小體串珠結構,其直徑10納米。然後按每圈6個核小體為單位盤繞成直徑30納米的螺線管。由螺線管形成DNA複製環,每18個複製環呈放射狀平面排列,結合在核基質上形成微帶。微帶是染色體高級結構的單位,大約10個微帶沿縱軸構建成子染色體。
功能
如果說細胞核是細胞遺傳與代謝的調控中心,那么這箇中心的最重要成員便是染色質。幾乎所有細胞生命活動都要從染色質開始。我們知道細胞的成長、分裂甚至衰老與死亡都是受基因控制的,而細胞內基因存在與發揮功能的結構基礎是染色質。與基因組直接相關的細胞活動都是在染色質水平進行的,如DNA複製、基因轉錄、同源重組、DNA修復,包括轉錄耦聯的修復以及DNA和組蛋白的各種修飾。這些修飾包括甲基化、乙醯化、磷酸化、亞硝基化和泛素化等。
真核生物的基因組都是在細胞核的三維空間中發揮功能,如基因組的複製、DNA 突變、DNA 修復、基因的轉錄和調控、長鏈非編碼 RNA 的傳播和胚胎髮育等。
組成物質介紹
 DNA
DNA細胞中編碼和控制的信息是與DNA分子緊密地聯繫在一起的。DNA與染色質有著重大聯繫。DNA是一種高分子聚合物,即由重複單位構成的大分子。每一單位都由三種較小分子組成,它們彼此結合形成核苷酸。鹼基共有四種:胸腺嘧啶(T),胞嘧啶(C),腺嘌呤(A),鳥嘌呤(G)。人的鹼基比例A:T:G:C是29.2:29.4:21.0:20.4,A+T/C+G比例為1.53。在多數來源DNA中,嘌呤的克分子數等於嘧啶的克分子數,即A+G=T+C,A=T,C=G。DNA分子由兩條螺線纏繞的分子鏈組成。鏈由糖和磷酸殘基交錯連線形成。每條鏈都有一個糖-磷酸主鏈和由糖部分向內突出的那些鹼基。在DNA分子中各個鹼基排列成對,DNA分子整體呈雙螺鏇形式。脫氧核糖與鹼基的特定碳原子連線,大分子中相鄰的兩個糖分子通過與磷酸形成的磷酸酯鍵彼此連線。鹼基藉助於氫鍵相互連線,使整個分子保持穩定。鹼基配對是嚴格互補的:A與T配對,C與G配對,DNA分子的兩條鏈在整個分子長度內是彼此互補的。
在人的細胞中,DNA是位於與膜相連的細胞核內,而蛋白質的合成則在細胞質里進行。細胞直接讀出的信息是編碼RNA分子上的,細胞不能直接閱讀染色體DNA上的信息。DNA中的胸腺嘧啶在RNA中由極為相似的分子尿嘧啶代替。RNA是單鏈的,與其模板DNA雙螺鏇中的一條多核苷酸鏈互補。這些信息最終被編譯成特定的結構蛋白或酶蛋白。
DNA是一種具有多種功能的分子。它能夠通過複製過程產生自己,細胞每分裂一次DNA便複製一次。它通過轉錄過程製造三種RNA:信使RNA(mRNA),轉移RNA(tRNA)和核糖體RNA(rRNA)。這三種RNA在細胞質合成蛋白質的一系列過程中行使不同的功能。信使RNA由DNA模板產生,是按一定順序排列的三聯體密碼子,因其是被讀出的信息而得名。密碼是一種三聯體,任一mRNA中核苷酸的數目至少應是所要合成的蛋白質中胺基酸數目的三倍。tRNA由好多種,是一類三葉草形狀的分子。tRNA成分中含有一些異常核苷酸,游離端的末端是核苷酸-CCA,中葉的頂部是三個核苷酸一組,構成反密碼子(可以用互補方式與mRNA三聯體密碼子的三個核苷酸配對),是mRNA編碼信息的讀者。
為了進行轉錄,DNA的雙螺鏇在RNA聚合酶的作用下被拆開。RNA聚合酶結合於雙鏈DNA,使它解鏇和拆開;同時形成RNA,RNA隨RNA聚合酶沿DNA的移動而增加長度。當mRNA從DNA中釋放出來,由細胞核進入細胞質,附著於一個或幾個核糖體上,其信息由相應的tRNA讀出。然後,附著於tRNA-CCA末端的胺基酸彼此連線,形成不斷加長的蛋白質。當tRNA到達終止密碼子(uAA, uAG, uGA)時,這一過程便告結束,合成的蛋白質被釋放出來,作為酶或膜以及其他細胞器的結構分子而供細胞使用。
 蛋白質
蛋白質蛋白質是荷蘭科學家格里特在1838年發現的。他觀察到有生命的東西離開了蛋白質就不能生存。蛋白質是生物體內一種極重要的高分子有機物,占人體乾重的54%。蛋白質主要由胺基酸組成,因胺基酸的組合排列不同而組成各種類型的蛋白質。人體中估計有10萬種以上的蛋白質。生命是物質運動的高級形式,這種運動方式是通過蛋白質來實現的,所以蛋白質有極其重要的生物學意義。人體的生長、發育、運動、遺傳、繁殖等一切生命活動都離不開蛋白質。生命運動需要蛋白質,也離不開蛋白質。人體內的一些生理活性物質如胺類、神經遞質、多肽類激素、抗體、酶、核蛋白以及細胞膜上、血液中起“載體”作用的蛋白都離不開蛋白質,它對調節生理功能,維持新陳代謝起著極其重要的作用。人體運動系統中肌肉的成分以及肌肉在收縮、作功、完成動作過程中的代謝無不與蛋白質有關,離開了蛋白質,體育鍛鍊就無從談起。在生物學中,蛋白質被解釋為是由胺基酸借肽鍵聯接起來形成的多肽,然後由多肽連線起來形成的物質。通俗易懂些說,它就是構成人體組織器官的支架和主要物質,在人體生命活動中,起著重要作用,可以說沒有蛋白質就沒有生命活動的存在。每天的飲食中蛋白質主要存在於瘦肉、蛋類、豆類及魚類中。蛋白質缺乏:成年人:肌肉消瘦、肌體免疫力下降、貧血,嚴重者將產生水腫。未成年人:生長發育停滯、貧血、智力發育差,視覺差。蛋白質過量:蛋白質在體內不能貯存,多了肌體無法吸收,過量攝入蛋白質,將會因代謝障礙產生蛋白質中毒甚至於死亡。
對比介紹
 染色質
染色質染色質、染色體和染色單體的區別
(1)、染色質和染色體的主要成分都是DNA和蛋白質,它們之間的不同,不過是同一物質在細胞分裂間期和分裂期的不同形態表現而已。
染色質出現於間期,呈絲狀。它們在核內的螺鏇程度不一,螺鏇緊密的部分,染色較深,有的螺旋鬆疏染色較淺,染色質在光鏡下呈現顆粒狀,不均勻地分布於細胞核中。細胞分裂時染色質細絲高度螺鏇化形成較粗的柱狀和桿狀等不同的形狀。不同生物的染色體(習慣不稱染色質)數目、形態不同,具有種的特異性,而且比較恆定。
(2)、每個染色體一般具有兩個臂或一個臂,兩臂之間有著絲點(是紡綞絲附著的地方)。細胞分裂間期由於染色體(習慣不稱染色質)複製形成由一個共同著絲點連在一起的兩個染色單體被稱為姐妹染色單體,這時的染色體仍為一條染色體。當細胞進入細胞有絲分裂後期,著絲點一分為二,姐妹染色單體也隨著分開,各有了自己的著絲點,這時就不再是染色單體而叫染色體了,隨之染色體數目加倍,染色單體消失。
染色體的組成
 染色體
染色體一個染色體一般呈棍棒狀,包含一個著絲點(c)和兩個臂(a、b)。著絲點是紡錘絲附著的地方,少數染色體的著絲點位於一端。一個染色體只有一個著絲點。因此,對染色體計數時就是看著絲點的數目。
染色體形態
在細胞周期中,染色體的形態有兩種,並且通過一定的方式相互轉化。A是通常所說的一個染色體。B是經過複製的染色體,包含兩個姐妹染色單體,兩個姐妹染色單體是完全相同的,其含有的物質也與A完全相同。B的著絲點分裂後,就變成了兩個完全相同的染色體,稱之為姐妹染色體。也就是說,染色體複製後至著絲點分裂之前,染色體的個數不變,但包含有染色單體,也僅在這一段時間內有染色單體。
染色體含分子數
A的一個染色體上有一個DNA分子,而B的染色體中含2個DNA分子,分別位於2個染色單體上。隨著著絲點分裂,B形成了C中的2個染色體,因而每個染色體只含一個DNA分子。
DNA分子數
要計算細胞中染色體上的DNA分子數:有染色單體時,DNA分子數=染色單體數,沒有染色單體時,DNA分子數=染色體數。
種類介紹
染色質著重介紹性染色質
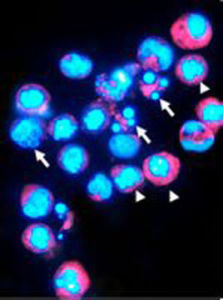
染色體在細胞周期的間期時DNA的螺鏇結構鬆散,呈網狀或斑塊狀不定形物,即染色質。以濃集狀態存在者,稱異染色質(1~eterochromatin);以分散狀態存在者,稱常染色質(euchromatin)。常染色質染色較淺且均勻,異染色質染色深。性染色質與性染色體(x染色體和Y染色體)有關,稱x染色質和Y染色質。
x染色質曾稱巴氏小體或X小體,為緊貼細胞核膜內面的團塊狀結構,直徑約1um,染色程度較其他染色質深。其形態不一,常呈三角、半圓、平凸或球形。利用放射自顯影技術的研究發現,女性的兩條x染色體中有一條DNA複製延遲,稱遲複製x。遲複製的x染色體在間期時表現為x染色質。當細胞內有一條以上x染色體時,在間期時除一條x染色體外,其餘的x染色體均表現為x染色質,因此間期細胞核中的x染色質數目等於x染色體數減去1。當x染色體結構異常時,x染色質的形態也會有相應的改變。如x等臂染色體時,出現大的x染色質,雙著絲粒x染色體時,出現雙葉或大的x染色質。
Y染色質又稱Y小體或螢光小體。Y染色體用螢光染料染色後,呈亮暗不一的螢光帶,在Y染色體長臂的遠側段呈明亮的螢光區。在問期時Y染色體長臂遠側段的強螢光特性仍然存在,經螢光染色後,呈強螢光亮點,直徑為0.25—0.3um,位於細胞核內的任何部位。
