
原核細胞的定義
原核細胞(prokaryotic cell)是組成原核生物的細胞。這類細胞主要特徵是沒有明顯可見的細胞核, 同時也沒有核膜和核仁, 只有擬核,進化地位較低。
 原核細胞
原核細胞原核細胞簡介
原核細胞(prokaryotic cell)沒有核膜,遺傳物質集中在一沒有明確界限的低電子密度區。DNA為裸露的環狀分子,通常沒有結合蛋白,環的直徑約為2.5nm,周長約幾十納米。沒有恆定的內膜系統,核糖體為70S型,原核細胞構成的生物稱為原核生物(prokaryote),均為單細胞生物,通常稱為細菌(bacterium)原核細胞 procaryotic/prokaryotic cell 指沒有核膜且不進行有絲分裂、減數分裂、無絲分裂的細胞。這種細胞不發生原生質流動,觀察不到變形蟲樣運動。鞭毛(flagellum)呈單一的結構。光合作用、氧化磷酸化在細胞膜進行,沒有(chloroplast)、線粒體(mitochondrion)等細胞器(organelles)的分化,只有核糖體。由這種細胞構成的生物,稱為原核生物,它包括所有的細菌和藍藻類。即構成細菌和藍藻等低等生物體的細胞。它沒有真正的細胞核(nucleus),只有原核或擬核,所含的一個基因帶(或染色體),是環狀雙股單一順序的脫氧核糖核酸分子(circular DNA),沒有組蛋白(histone)與之結合無核仁(nucleolus),缺乏核(nuclearenvelope)。外層原生質中有70 S核糖體與中間體,缺乏高爾基體(Golgi)、內質網(E.R.)、線粒體和中心體(centrosomes)等。轉錄和轉譯(transcription and translation)同時進行,四周質膜內含有呼吸酶。無有絲分裂(mitosis)和減數分裂(meiosis),脫氧核糖核酸(DNA)複製後,細胞隨即分裂為二結構含有capsule,murein cell wall,cell surface membrane,circular DNA, mesosome, thykoloid, energy storage(e.g.glycogen), ribosome, flagellum, pilli 等等 。
原核細胞主要生物種類
原核細胞構成的生物稱為原核生物(prokaryote),均為單細胞生物,通常稱為細菌(bacterium)。
一、細菌細菌是在自然界分布最廣、個體數量最多的有機體,是大自然物質循環的主要參與者。細菌主要由細胞壁、細胞膜、細胞質、核質體等部分構成,有的細菌還有夾膜、鞭毛、菌毛等特殊結構。絕數細菌的直徑大小在0.5~5μm之間。可根據形狀三類,即:球菌、桿菌和螺鏇菌。
 原核細胞
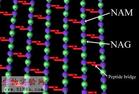
原核細胞(一)細胞壁細胞壁厚度因細菌不同而異,一般為15-30nm。主要成分是肽聚糖,由N-乙醯葡糖胺和N-乙醯胞壁酸構成雙糖單元,以β(1-4)糖苷鍵連線成大分子。N-乙醯胞壁酸分子上有四肽側鏈,相鄰聚糖纖維之間的短肽通過肽橋(革蘭氏陽性菌)或肽鍵(革蘭氏陰性菌)橋接起來,形成了肽聚糖片層,像膠合板一樣,粘合成多層肽聚糖中的多糖鏈在各物種中都一樣,而橫向短肽鏈卻有種間差異。革蘭氏陽性菌細胞壁厚約20~80nm,有15-50層肽聚糖片層,每層厚1nm,含20-40%的磷壁酸(teichoic acid),有的還具有少量蛋白質。革蘭氏陰性菌細胞壁厚約10nm,僅2-3層肽聚糖,其他成分較為複雜,由外向內依次為脂多糖、細菌外膜和脂蛋白。此外,外膜與細胞之間還有間隙。肽聚糖是革蘭陽性菌細胞壁的主要成分,凡能破壞肽聚糖結構或抑制其合成的物質,都有抑菌或殺菌作用。如溶菌酶是N-乙醯胞壁酸酶,青黴素抑制轉肽酶的活性,抑制肽橋形成細菌細胞壁的功能包括:保持細胞外形;抑制機械和滲透損傷(革蘭氏陽性菌的細胞壁能耐受20kg/cm2的壓力);介導細胞間相互作用(侵入宿主);防止大分子入侵;協助細胞運動和分裂脫壁的細胞稱為細菌原生質體(bacterial protoplast)或球狀體(spheroplast,因脫壁不完全),脫壁後的細菌原生質體,生存和活動能力大大降低。
(二)細胞膜是典型的單位膜結構,厚約8~10nm,外側緊貼細胞壁,某些革蘭氏陰性菌還具有細胞外膜。通常不形成內膜系統,除核糖體外,沒有其它類似真核細胞的細胞器,呼吸和光合作用的電子傳遞鏈位於細胞膜上。某些行光合作用的原核生物(藍細菌和紫細菌),質膜內褶形成結合有色素的內膜,與捕光反應有關。某些革蘭氏陽性細菌質膜內褶形成小管狀結構,稱為中膜體(mesosome)或間體,中膜體擴大了細胞膜的表面積,提高了代謝效率,有擬線粒體(Chondroid)之稱,此外還可能與DNA的複製有關。
(三)細胞質與核質體細菌和其它原核生物一樣,沒有核膜,DNA集中在細胞質中的低電子密度區,稱核區或核質體(nuclear body)。細菌一般具有1-4個核質體,多的可達20餘個。核質體是環狀的雙鏈DNA分子,所含的遺傳信息量可編碼2000~3000種蛋白質,空間構建十分精簡,沒有內含子。由於沒有核膜,因此DNA的複製、RNA的轉錄與蛋白的質合成可同時進行,而不像真核細胞那樣些生化反應在時間和空間上是嚴格分隔開來的每個細菌細胞約含5000~50000個核糖體,部分附著在細胞膜內側,大部分游離於細胞質中。細菌核糖體的沉降係數為70S,由大亞單位(50S)與小亞單位(30S)組成,大亞單位含有23SrRNA,5SrRNA與30多種蛋白質,小亞單位含有16SrRNA與20多種蛋白質。30S的小亞單位對四環素與鏈黴素很敏感,50S的大亞單位對紅黴素與氯黴素很敏感。
細菌核區DNA以外的,可進行自主複製的遺傳因子,稱為質粒(plasmid)。質粒是裸露的環狀雙鏈DNA分子,所含遺傳信息量為2~200個基因,能進行自我複製,有時能整合到核DNA中去。質粒DNA在遺傳工程研究中很重要,常用作基因重組與基因轉移的載體胞質顆粒是細胞質中的顆粒,起暫時貯存營養物質的作用,包括多糖、脂類、多磷酸鹽等。
(四)其他結構許多細菌的最外表還覆蓋著一層多糖類物質,邊界明顯的稱為莢膜,如肺炎球菌,邊界不明顯的稱為粘液層(slime layer),如葡萄球菌。莢膜對細菌的生存具有重要意義,細菌不僅可利用莢膜抵禦不良環境;保護自身不受白細胞吞噬;而且能有選擇地粘附到特定細胞的表面上,表現出對靶細胞的專一攻擊能力。例如,傷寒沙門桿菌能專一性地侵犯腸道淋巴組織。細菌莢膜的纖絲還能把細菌分泌的消化酶貯存起來,以備攻擊靶細胞之用。鞭毛是某些細菌的運動器官,由一種稱為鞭毛蛋白(flagellin)的彈性蛋白構成,結構上不同於真核生物的鞭毛。細菌可以通過調整鞭毛鏇轉的方向(順和逆時針)來改變運動狀態。菌毛是菌體表面極其的蛋白纖細,須用電鏡觀察。特點是:細、短、直、硬、多,菌毛與細菌運動無關,根據形態、結構和功能,可分為普通菌毛和性菌毛兩類。前者與細菌吸附和侵染宿主有關,後者為中空管子,與傳遞遺傳物質有關。
(五)繁殖細菌一二分裂的方式繁殖,某些細菌處於不利的環境,或耗盡營養時,形成內生孢子,又稱芽孢,是對不良環境有強抵抗力的休眠體,由於芽胞在細菌細胞內形成,故常稱為內生孢子。
芽孢的生命力非常頑強,有些湖底沉積土中的芽抱桿茵經500-1000年後仍有活力,肉毒梭菌的芽孢在pH 7.0時能耐受100℃煮沸5-9.5小時。芽孢由內及外有以下幾部分組成:
1.芽孢原生質(spore protoplast,核心core):含濃縮的原生質。
2.內膜(inner membrane):由原來繁殖型細菌的細胞膜形成,包圍芽孢原生質。
3.芽孢壁(spore wall):由繁殖型細菌的肽聚糖組成,包圍內膜。發芽後成為細菌的細胞壁。
4.皮質(cortex):是芽孢包膜中最厚的一層,由肽聚糖組成,但結構不同於細胞壁的肽聚糖,交聯少,多糖支架中為胞壁酐而不是胞壁酸,四肽側鏈由L-Ala組成。
5.外膜(outer membrane):也是由細菌細胞膜形成的。
 原核細胞
原核細胞6.外殼(coat):芽孢殼,質地堅韌緻密,由類角蛋白組成(keratinlike protein),含有大量二硫鍵,具疏水性特徵。
7.外壁(exosporium):芽孢外衣,是芽孢的最外層,由脂蛋白及碳水化合物(糖類)組成,結構疏鬆。
二、支原體支原體(mycoplasma)的大小通常為0.2~0.3μm,可通過濾菌器。無細胞壁,不能維持固定的形態而呈現多形性。細胞膜中膽固醇含量較多,約占36%,這對保持細胞膜的完整性是必需的,凡能作用於膽固醇的物質(如二性黴素B、皂素等)均可引起支原體膜的破壞而使支原體死亡。
支原體基因組為一環狀雙鏈DNA,分子量小(僅有大腸桿菌的五分之一),合成與代謝很有限。肺炎支原體的一端有一種特殊的末端結構(terminal structure),能使支原體粘附於呼吸道黏膜上皮細胞表面,與致病性有關。
三、衣原體和立克次氏體衣原體(Chlamydia)很小,直徑200nm-500nm,能通過細菌濾膜。立克次氏體(Rickettsia)略大,大多不能通過濾菌膜。它們都有DNA和RNA,有革蘭氏陰性細菌特徵的含肽聚糖的細胞壁,但酶系統不完全,必須在寄主細胞內生活,有攝能寄生物(energe parasite)之稱砂眼是衣原體引起的,由於能形成包含體,起初被認為是大型病毒,1956年,我國著名微生物學家湯飛凡及其助手張曉樓等人首次分離到沙眼的病原體衣原體生活史特殊,具有感染力的個體稱為原體(elementory body),體積小,有堅韌的細胞壁。在宿主細胞內,原體逐漸伸長,形成無感染力的個體,稱作始體(initial body),是一種薄壁的球狀細胞,體積較大,通過二等分裂的方式在宿主細胞內形成一個微菌落,隨後大量的子細胞有分化為具有感染能力的原體立克次氏體也是專性細胞內寄生的,主要寄生於節肢動物,有的會通過蚤、虱、蜱、蟎傳入人體、如斑疹傷寒、戰壕熱。美國醫生H.T.Richetts 1909年首次發現它是落基山斑疹傷寒的病原體,並於1910年犧牲於此病,故後人稱這類病原體為立克次氏體。與衣原體的不同處在於其細胞較大,無濾過性,合成能力較強,且不形成包涵體。
四、藍藻藍藻又稱藍細菌(cyanobacterium),能進行與高等植物類似的光合作用(以水為電子供體,放出O2),與光合細菌的光合作用的機制不一樣,因此被認為是最簡單的植物。藍藻沒有葉綠體,僅有十分簡單的光合作用結構裝置。藍藻細胞遺傳信息載體與其它原核細胞一樣,是一個環狀DNA分子,但遺傳信息量很大,可與高等植物相比。藍藻細胞的體積比其它原核細胞大得多,直徑一般在士10um,甚至可達70μm(顫藻)。藍藻屬單細胞生物,有些藍藻經常以絲狀的細胞群體存在,如:屬藍藻門念珠藻類的髮菜(nostoc commune var.flagtlliforme)就是藍藻的絲狀體;做綠肥的紅萍實際上是一種固氮藍藻與水生蕨類滿江紅的共生體。
 原核細胞
原核細胞細胞結構與生物系統
傳統分類法根據生物的營養方式、運動能力和細胞結構的特點,把生物劃分為動物界和植物界。植物細胞的主要特徵是具有硬的細胞壁和進行光合作用的葉綠體。按傳統分類系統,雖然大多數生物種容易歸類,可是對某些生物來說卻遇到了分類上的困難,例如眼蟲(Euglena)是一種單細胞生物,含有葉綠體,卻不具有細胞壁;細菌和真菌則有細胞壁而無葉綠體;支原體既無葉綠體也無細胞壁,古細菌既有原核生物的特徵也具有真核生物的特徵。這些生物按照傳統分類法進行分類顯然就要遇到困難1977年C. Woese根據對16SrRNA核苷酸順序的同源性比較,提出將生命劃分為三界,即:真細菌(Eubacteria)、真核生物Eucaryotes、古細菌(Archaes)。1996年Bult領導的研究小組在Science上發表了詹氏甲烷球菌(Methanococcus jannaschii)的全基因組序列,進一步證明它既不是典型的細菌也不是典型的真核生物,而是介於兩者之間的生命體,即生命的第三形式。
 原核細胞系統
原核細胞系統細菌
細菌是在自然界分布最廣、個體數量最多的有機體,是大自然物質循環的主要參與者。細菌主要由細胞壁、細胞膜、細胞質、核質體等部分組成,有的細菌還有莢膜、鞭毛、菌毛等特殊結構。絕大多數細菌的直徑大小在0.5~5μm之間。可根據形狀分成三類:球菌、桿菌和螺鏇菌。桿菌螺鏇菌一、細胞壁
細胞壁厚度因細菌不同而異,一般為15-30nm。主要成分是肽聚糖,由N-乙醯葡糖胺和N-乙醯胞壁酸構成雙糖單元,以β(1-4)糖苷鍵連線成大分子。N-乙醯胞壁酸分子上有四肽側鏈,相鄰聚糖纖維之間的短肽通過肽橋(革蘭氏陽性菌)或肽鍵(革蘭氏陰性菌)橋接起來,形成了肽聚糖片層,像膠合板一樣,粘合成多層。肽聚糖中的多糖鏈在各物種中都一樣,而橫向短肽鏈卻有物種間的差異。革蘭氏陽性菌細胞壁厚約20~80nm,有15-50層肽聚糖片層,每層厚1nm,含20-40%的磷壁酸(teichoicacid),有的還含有少量蛋白質。革蘭氏陰性菌細胞壁厚約10nm,僅2-3層肽聚糖,其他成分較為複雜,由外向內依次為脂多糖、細菌外膜和脂蛋白。此外,外膜與細胞之間還有間隙。
肽聚糖是革蘭陽性菌細胞壁的主要成分,凡能破壞肽聚糖結構或抑制其合成的物質,都有抑菌或殺菌作用。如溶菌酶是N-乙醯胞壁酸酶,青黴素抑制轉肽酶的活性和肽橋的形成。
細菌細胞壁的功能包括:保持細胞外形;抑制機械和滲透損傷(革蘭氏陽性菌的細胞壁能耐受20kg/c㎡的壓力);介導細胞間相互作用(侵入宿主);防止大分子入侵;協助細胞運動和分裂。
脫壁的細胞稱為細菌原生質體(bacterialprotoplast)或球狀體(spheroplast),因脫壁不完全,脫壁後的細菌原生質體,生存和活動能力大大降低。
細胞膜
是典型的單位膜結構,厚約8~10nm,外側緊貼細胞壁,某些革蘭氏陰性菌還具有細胞外膜。細胞膜有多方面的重要功能,它與細胞的物質交換,細胞識別,分泌,排泄,免疫等都有密切的關係。通常不形成內膜系統,除核糖體外,沒有其它類似真核細胞的細胞器,呼吸和光合作用的電子傳遞鏈位於細胞膜上。某些行光合作用的原核生物(藍細菌和紫細菌),質膜內褶形成結合有色素的內膜,與捕光反應有關。某些革蘭氏陽性細菌質膜內褶形成小管狀結構,稱為中膜體(mesosome)或間體,中膜體擴大了細胞膜的表面積,提高了代謝效率,有擬線粒體(Chondroid)之稱,此外還可能與DNA的複製有關。
細菌的基本結構細胞質與核質體
細菌和其它原核生物一樣,沒有核膜,DNA集中在細胞質中的低電子密度區,稱核區或核質體(nuclearbody)。細菌一般具有1-4個核質體,多的可達20餘個。核質體是環狀的雙鏈DNA分子,所含的遺傳信息量可編碼2000~3000種蛋白質,空間構建十分精簡,沒有內含子。由於沒有核膜,因此DNA的複製、RNA的轉錄與蛋白的質合成可同時進行,而不像真核細胞那樣在生化反應在時間和空間上是嚴格分隔開來的。
每個細菌細胞約含5000~50000個核糖體,部分附著在細胞膜內側,大部分游離於細胞質中。細菌核糖體的沉降係數為70S,由大亞單位(50S)與小亞單位(30S)組成,大亞單位含有23SrRNA與30多種蛋白質,小亞單位含有16SrRNA與20多種蛋白質。30S的小亞單位對四環素與鏈黴素很敏感,50S的大亞單位對紅黴素與氯黴素很敏感。
細菌核區DNA以外的,可進行自主複製的遺傳因子,稱為質粒(plasmid)。質粒是裸露的環狀雙鏈DNA分子,所含遺傳信息量為2~200個基因,能進行自我複製,有時能整合到核DNA中去。質粒DNA在遺傳工程研究中很重要,常用作基因重組與基因轉移的載體。
胞質顆粒是細胞質中的顆粒,起暫時貯存營養物質的作用,包括多糖、脂類、多磷酸鹽等。
其他結構
許多細菌的最外表還覆蓋著一層多糖類物質,其中邊界明顯的稱為莢膜,如肺炎球菌,邊界不明顯的稱為粘液層(slimelayer),如葡萄球菌。莢膜對細菌的生存具有重要意義,細菌不僅可利用莢膜抵禦不良環境;保護自身不受白細胞吞噬;而且能有選擇地粘附到特定細胞的表面上,表現出對靶細胞的專一攻擊能力。例如,傷寒沙門桿菌能專一性地侵犯腸道淋巴組織。細菌莢膜的纖絲還能把細菌分泌的消化酶貯存起來,以備攻擊靶細胞之需。
鞭毛是某些細菌的運動器官,由一種稱為鞭毛蛋白(flagellin)的彈性蛋白構成,結構上不同於真核生物的鞭毛。細菌可以通過調整鞭毛鏇轉的方向(順時針和逆時針)來改變運動狀態。
菌毛是菌體表面極其細的蛋白纖維,須用電鏡觀察,特點是:細、短、直、硬、多。菌毛與細菌運動無關,根據形態、結構和功能,可分為普通菌毛和性菌毛兩類。前者與細菌吸附和侵染宿主有關,後者為中空管子,與傳遞遺傳物質有關。
繁殖
細菌以一分為二的方式繁殖,某些細菌處於不利的環境,或耗盡營養時,形成內生孢子,又稱芽孢,是對不良環境有強抵抗力的休眠體,由於芽胞在細菌細胞內形成,故常稱為內生孢子。
芽孢的生命力非常頑強,有些湖底沉積土中的芽孢桿菌經500-1000年後仍有活力,肉毒梭菌的芽孢在pH7.0時能耐受100℃煮沸5-9.5小時。芽孢由內及外有以下幾部分組成:
1.芽孢原生質(sporeprotoplast):核心(core)含濃縮的原生質。
2.內膜(innermembrane):由原來繁殖型細菌的細胞膜形成,包圍芽孢原生質。
3.芽孢壁(sporewall):由繁殖型細菌的肽聚糖組成,包圍內膜。發芽後成為細菌的細胞壁。
4.皮質(cortex):是芽孢包膜中最厚的一層,由肽聚糖組成,但結構不同於細胞壁的肽聚糖,交聯少,多糖支架中為胞壁酐而不是胞壁酸,四肽側鏈由L-Ala組成。
5.外膜(outermembrane):也是由細菌細胞膜形成的。
6.外殼qiao(coat):芽孢外殼,質地堅韌緻密,由類角蛋白(keratinlikeprotein)組成,含有大量二硫健,具有疏水性特徵。
7.外壁(exosporium):芽孢外衣,是芽孢的最外層,由脂蛋白及碳水化合物(糖類)組成,結構疏鬆。
放線菌
放線菌是具有菌絲、以孢子進行繁殖、革蘭氏染色陽性的一類原核微生物。因其具有分枝狀菌絲、菌落形態與黴菌相似,過去曾認為放線菌是"介於細菌與真菌之間的微生物"。然而,用近代生物學技術所進行的研究結果表明,放線菌實際上是屬於細菌範疇內的原核微生物,只不過其細胞形態為分枝狀菌絲。從系統發育上看,放線菌(除高溫放線菌外)與全部G+細菌一起同屬於這一大分支中的高G+C/mol%群。藍藻
藍藻又稱藍細菌(cyanobacterium),能進行與高等植物類似的光合作用藍藻(以水為電子供體,放出O2),與光合細菌的光合作用的機制不一樣,因此被認為是最簡單的植物。藍藻沒有葉綠體,但含有藻藍素(呈藍色,但含量少)和葉綠素(呈綠色並且含量多),是能夠進行光合作用的自養生物。細菌中絕大多數種類是營腐生或者寄生生活的一種生物。藍藻和細菌中,都沒有成型的細胞核。藍藻細胞遺傳信息載體與其它原核細胞一樣,是一個環狀DNA分子,但遺傳信息量很大,可與高等植物相比。因為藍藻細胞的體積比其它原核細胞大得多,直徑一般在10um,甚至可達70μm(顫藻),所以肉眼不能看清藍藻細胞。藍藻屬單細胞生物,有些藍藻經常以絲狀的細胞群體存在,如:屬藍藻門念珠藻類的髮菜(nostoccommunevar.flagtlliforme)就是藍藻的絲狀體;髮菜生長在西北草地和荒漠,因為和“發財”諧音,所以被人爭相食用,現已被我國列為保護生物;做綠肥的紅萍實際上是一種固氮藍藻與水生蕨類滿江紅的共生體。藍球藻,念珠藻,顫藻,髮菜(細胞群體呈黑藍色)均屬於藍藻,所以藍藻並不都是呈現綠色的。支原體
膽固醇支原體(mycoplasma)的大小通常為0.2~0.3μm,是現認為最小的細胞,可通過濾菌器。無細胞壁,支原體不能維持固定的形態而呈現多形性。細胞膜中膽固醇含量較多,約占36%,這對保持細胞膜的完整性是必需的,凡能作用於膽固醇的物質(如二性黴素B、皂素等)均可引起支原體膜的破壞而使支原體死亡。支原體基因組為一環狀雙鏈DNA,分子量小(僅有大腸桿菌的五分之一),合成與代謝很有限。肺炎支原體的一端有一種特殊的末端結構(terminalstructure),能使支原體粘附於呼吸道黏膜上皮細胞表面,與致病性有關。
衣原體和立克次氏體
衣原體(Chlamydia)很小,直徑200nm-500nm,能通過細菌濾膜。立克次氏體(Rickettsia)略大,大多不能通過濾菌膜。它們都有DNA和RNA,有革蘭氏陰性細菌特徵的含肽聚糖的細胞壁,但酶系統不完全,必須在寄主細胞內生活,有攝能寄生物(energeparasite)之稱。砂眼是衣原體引起的,由於能形成包含體,起初被認為是大型病毒,1956年,中國著名微生物學家湯飛凡及其助手張曉樓等人首次分離到沙眼的病原體。
衣原體生活史特殊,具有感染力的個體稱為原體(elementorybody),體積小,有堅韌的細胞壁。在宿主細胞內,原體逐漸伸長,形成無感染力的個體,稱作始體(initialbody),是一種薄壁的球狀細胞,體積較大,通過二等分裂的方式在宿主細胞內形成一個微菌落,隨後大量的子細胞有分化為具有感染能力的原體。立克次氏體立克次氏體也是專性細胞內寄生的,
主要寄生於節肢動物,有的會通過蚤、虱、蜱、蟎傳入人體、如斑疹傷寒、戰壕熱。美國醫生H.T.Richetts
1909年首次發現它是落基山斑疹傷寒的病原體,並於1910年犧牲於此病,故後人稱這類病原體為立克次氏體。與衣原體的不同處在於其細胞較大,無濾過
性,合成能力較強,且不形成包涵體。
細胞結構與生物系統
細菌傳統分類法根據生物的營養方式、運動能力和細胞結構的特點,把生物劃分為動物界和植物界。植物細胞的主要特徵是具有硬的細胞壁和進行光合作用的葉綠體。按傳統分類系統,雖然大多數生物種容易歸類,可是對某些生物來說卻遇到了分類上的困難,例如眼蟲(Euglena)是一種單細胞生物,含有葉綠體,卻不具有細胞壁;細菌和真菌則有細胞壁而無葉綠體;支原體既無葉綠體也無細胞壁,古細菌既有原核生物的特徵也具有真核生物的特徵。這些生物按照傳統分類法進行分類顯然就要遇到困難。1977年C.Woese根據對16SrRNA核苷酸順序的同源性比較,提出將生命劃分為三界,即:真細菌(Eubacteria)、真核生物Eucaryotes、古細菌(Archaes)。1996年Bult領導的研究小組在Science上發表了詹氏甲烷球菌(Methanococcusjannaschii)的全基因組序列,進一步證明它既不是典型的細菌也不是典型的真核生物,而是介於兩者之間的生命體,即生命的第三形式。
基因結構
原核生物的基因結構多數以操縱子形式存在,即完成同類功能的多個基因聚集在一起,處於同一個啟動子的調控之下,下游同時具有一個終止子。兩個基因之間存在長度不等的間隔序列,如與乳糖代謝有關酶的基因。在距轉錄起始點-35和-10(轉錄起始點上游的核苷酸序列為“-”,下游的核苷酸序列為“+”)附近的序列都有RNA聚合酶識別的信號。RNA聚合酶先與-35附近的序列(稱為Pribnow框)結合,然後才與
-10附近的序列(稱為Sextama框)結合。至於RNA聚合酶是如何從一個位置轉到另一個位置的,目前尚不清楚。RNA聚合酶一旦與-10附近序列結
合,就立即從識別位點上脫離下來,DNA雙鏈解開,轉錄開始。除啟動子外,往往還有一些調控轉錄的其他因子,如調節基因和操縱基因。
原核生物基因轉錄終止之前同樣有一段迴文序列結構,稱為終止子,它的特殊的鹼基排列順序能夠阻礙RNA聚合酶的移動,並使其從DNA模板鏈上脫離下來。
原核細胞的基因結構
核生物的基因結構多數以操縱子形式存在(見課本第二節中的乳糖操縱子),即完成同類功能的多個基因聚集在一起,處於同一個啟動子的調控之下,下游同時具有一個終止子。兩個基因之間存在長度不等的間隔序列,如與乳糖代謝有關酶的基因。在距轉錄起始點-35和-10(轉錄起始點上游的核苷酸序列為“-”,下游的核苷酸序列為“+”)附近的序列都有RNA聚合酶識別的信號。RNA聚合酶先與-35附近的序列(稱為Pribn
 原核細胞結構
原核細胞結構