
生物起源
脊索動物無疑是由無脊椎動物起源的。而其中的棘皮動物和半索動物與脊索動物的親緣關係較近的觀點為大多數人所接受。
經推測,脊索動物的祖先可能類似於尾索動物的幼體,它向兩個方向發展,一是經過變態,成體為固者生活,具鰓裂作為取食和呼吸器官;另一個方向是幼體期延長並適應新的生活環境,不再變態,產生生殖腺並進行繁殖(即幼體性成熟),進而發展出新的一類動物,即具有脊索、背神經管和鰓裂的自由運動的脊索動物;之後分化為有頜類(魚類祖先)和無頜類。
形態特徵
共同特徵
脊索
位於消化道和神經管之間的一條棒狀結構,具有支持功能,所有脊索動物的胚胎期具有脊索,但在以後的生活中或終生保留(尾索動物亞門、頭索動物亞門),或退化並被脊柱(vertebral column)代替。
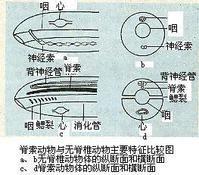 脊索動物門
脊索動物門 脊索來源於胚胎期的原腸背壁,即脊索中胚層。經加厚、分化、外突,最後脫離原腸而成脊索。脊索由富含液泡的脊索細胞組成,外面圍有脊索細胞所分泌而形成的結締組織性質的脊索鞘(notochordal sheath)。脊索鞘常包括內外兩層,分別為纖維組織鞘(fibrous sheath)和彈性組織鞘(elastic sheeth)。充滿液泡的脊索細胞由於產生膨壓,使整條脊索既具彈性,又有硬度, 從而起到骨骼的基本作用。低等脊索動物中,脊索終生存在或僅見於幼體時期。高等脊索動物只在胚胎期間出現脊索,發育完全時即被分節的骨質脊柱(vertebral column)所取代。組成脊索或脊柱等內骨骼(endoskeleton)的細胞,都能隨同動物體發育而不斷生長。而無脊椎動物則缺乏脊索或脊柱等內骨骼,通常僅身體表面被有幾丁質等外骨骼(exoskeleton)。脊索的出現在動物演化史上具有重要意義。表現在:
①脊索(以及脊柱)構成支撐軀體的主梁,是體重的受力者,也是內臟器官得到有力的支持和保護。
②運動肌肉獲得堅強的支點,在運動時不致由於肌肉的收縮而使軀體縮短或變形,因而向“大型化”發展。同時,脊索的中軸支撐作用也能使動物體更有效地完成定向運動,對於主動捕食及逃避敵害都更為準確、迅速。
③脊椎動物頭骨的形成、頜的出現以及椎管對中樞神經的保護,都是在此基礎上進一步完善化的發展。
脊髓(dorsal tubular nerve cord)
位於脊索背面中空管狀的中樞神經系統。脊椎動物神經管前端膨大成腦,腦後部分形成脊髓。
由胚體背中部的外胚層內陷形成。背神經管在高等種類中前、後分化為腦和脊髓。神經管腔(neurocoele)在腦內形成腦室(cerebral ventricle),在脊髓中成為中央管(centralcanal)。無脊椎動物神經系統的中樞部分為一條實性的腹神經索(ventral nerve cord),位於消化道的腹面。
鰓裂
 脊索動物門
脊索動物門 咽部兩側一系列左右成對排列的裂縫,直接或間接與外界相通。低等脊索動物及魚類的鰓裂終生存在,其他脊椎動物僅在胚胎期有鰓裂。來源於外胚層和內胚層
低等脊索動物在消化道前端的咽部兩側有一系列左右成對排列、數目不等的裂孔,直接開口於體表或以一個共同的開口間接地與外界相通,這些裂孔就是咽鰓裂。低等水棲脊索動物的鰓裂終生存在並附生著布滿血管的鰓,作為呼吸器官,陸棲高等脊索動物僅在胚胎期或幼體期(如兩棲綱的蝌蚪)具有鰓裂,隨同發育成長最終完全消失。無脊椎動物的鰓不位於咽部,用作呼吸的器官有軟體動物的櫛鰓以及節肢動物的肢鰓、尾鰓、氣管等。
肛尾
尾在肛門後方。肛後尾在所有的脊索動物中出現,至少在胚胎期(如人類)出現。
以上的前3個特徵是脊索動物的主要3大特徵,此外的肛後尾及閉管式循環系統(尾索動物除外);心臟位於消化道腹面;胚胎期原腸胚的發育類型為原口;分節的肌節,附著在不分節的軀幹上等特徵為脊索動物的次要特徵。
無脊椎動物的肛孔常開口在軀幹部的末端。
循環系統
位於消化道的腹面,循環系統為閉管式。無脊椎動物的心臟及主動脈在消化道的背面,循環系統大多為開管式。
在上述特徵中,具有脊索、背神經管和咽鰓裂是區別脊索動物和無脊椎動物最主要的3個基本特徵。此外,脊索動物還有一些性狀同樣也見於高等無脊椎動物的,例如三胚層、後口、存在次級體腔、兩側對稱的體制、身體和某些器官的分節現象等。這些共同點表明脊索動物是由無脊椎動物進化而來的。
尾索動物
1、體壁:柄海鞘的體壁即是包藏內部器官的外套膜(mantle),外套膜除了表面一層外胚層的上皮細胞外,還參雜著來源於中胚層的肌肉纖維,以支配身體及出、入水孔的伸縮和開關。體壁能分泌一種化學成分類似植物纖維素的被囊素(tunicin),並由此形成包圍在動物體外的被囊,這就是被囊動物名稱的由來。在整個動物界中,體壁能分泌被囊素的動物,至今僅發現於尾索動物和少數原生動物。外套膜在入水孔和出水孔的邊緣處與被囊匯合,匯合處有環形括約肌控制管孔的啟閉。內部器官中只有咽的上緣及腹面的一部分與外套膜癒合。柄海鞘的被囊表面通常不易被其它動物所附著,但是同種個體卻能重疊附生,這對種群的繁衍顯然是有積極意義的。
2、消化和呼吸系統:入水孔的底部有口,通過四周長有觸手的緣膜就是寬大的咽,咽幾乎占據了身體的大半部(3/4),咽壁被許多細小的鰓裂所貫穿。從口進入咽內的水流經過鰓裂,到達圍著咽外的圍鰓腔中,然後 經出水孔排出。圍鰓腔是由身體表面陷入內部所形成的空腔,因其不斷擴大,從而將身體前部原有的體腔逐漸擠小,最終在咽部完全消失。由於鰓裂的間隔里分布著豐富的毛細血管,因此當水流攜帶著食物微粒通過鰓裂時就能進行氣體交換,完成呼吸作用。咽腔的內壁生有纖毛,其背、腹側的中央各有一溝狀結構,分別稱為背板(dorsal lamina或咽上溝epipharyngeal groove)和內柱(endostyle),溝內有腺細胞和纖毛細胞;背板和內柱上下相對,在咽的前端以圍咽溝(peripharyngeal groove)相連,腺細胞能分泌粘液,使沉入內柱的食物粘聚成團,由溝內的纖毛擺動,將食物團從內柱推向前行,經圍咽溝沿背板往後導入食道、胃及腸進行消化。腸開口於圍鰓腔,不能消化的殘渣通過圍鰓腔,隨水流經出水孔排出體外。
3、循環方式和排泄器官:心臟位於身體腹面靠近胃部的圍心腔(pericardial cavity)內,藉圍心膜的伸縮而搏動。心臟兩端各發出一條血管,前端一條為鰓血管,沿咽腹發出分支到鰓裂間的咽壁上;後端一條稱腸血管,分支到各內臟器官並注血進入器官組織的血竇之間,所以是開管式的血液循環。柄海鞘具有特殊的可逆式血液循環流向,即心臟收縮有周期性間歇,當它的前端連續搏動時,血液不斷地由鰓血管壓出至鰓部,接著心臟有短暫的停歇,容納鰓部的血液流回心臟,然後於其後端開始搏動,將血液注入腸血管而分布到內臟器官的組織間。因此,柄海鞘的血管既無動脈和靜脈之分,血液也無固定的單向流動方向,這種獨特的血液循環方式在動物界中是絕無僅有的。
柄海鞘無專門的排泄器官,僅在腸附近有一堆具排泄機能的細胞,稱為小腎囊(renal vesicles),其中常含尿酸結晶。
4、神經:柄海鞘的成體營固著生活,神經系統和感覺器官均甚退化,中樞神經只是一個沒有內腔的神經節(nervus ganglion),圓而堅硬,狀如小瘤,位於入水孔和出水孔之間的外套膜壁內,由此分出若干神經分支到身體各部,神經節旁有一無色透明而略為膨大的神經腺(neural gland,相當高等動物的腦下腺hypophy-sis)。無專門的感覺器官,僅於入水管孔、出水管孔的緣膜和外套膜上有少量散在的感覺細胞。
5、生殖系統:雌雄同體,生殖腺位於腸環間和外套膜內壁上。精巢大,呈分支狀,為乳白色顆粒狀小塊;卵巢長管狀,呈淡黃色,內含許多圓形的卵細胞;兩者緊貼重疊,分別以單根生殖導管(gonoduct)將成熟的性細胞輸入圍鰓腔,然後經出水管孔排至體外,或在圍鰓腔內與另一海鞘的生殖細胞相遇受精。
頭索動物
1、皮膚:皮膚薄而半透明,由單層柱形細胞的表皮和凍膠狀結締組織的真皮兩部分構成,表皮外覆有一層角皮層(cuticle)。表皮外在幼體期生有纖毛,成長後則消失殆盡。
2、骨骼:文昌魚尚未形成骨質的骨骼,主要是以縱貫全身的脊索作為支持動物體的中軸支架。脊索外圍有脊索鞘膜,並與背神經管的外膜、肌節之間的肌隔、皮下結締組織等連續。脊索細胞呈扁盤狀,其超顯微結構與雙殼類軟體動物的肌細胞比較相似,收縮時可增加脊索的硬度。此外,在口笠觸鬚、緣膜觸手、輪器內部也都有角質物支持,奇鰭和鰓裂的鰭條(fin rays)及鰓條(gill bar)由結締組織支持。
3、肌肉:文昌魚背部的肌肉厚實而腹部比較單薄,與無脊椎動物周身體壁厚薄均勻的情況不同。全身主要的肌肉是60多對按節排列於體側的較原始的"V"字形肌節(myomere),尖端朝前,肌節間被結締組織的肌隔(myocomma)所分開。兩側的肌節互不對稱,便於文昌魚在水平方向作彎曲運動。此外,還有分布在圍鰓腔腹面的橫肌和口緣膜上的括約肌等,控制圍鰓腔的排水及口孔的大小。
4、消化和呼吸器官:文昌魚靠輪器和咽部纖毛的擺動,使帶有食物微粒的水流經口入咽,食物被濾下留在咽內,而水則通過咽壁的鰓裂至圍鰓腔,然後由腹孔排出體外。作為收集食物和呼吸場所的咽部極度擴大,幾乎占據身體全長的1/2,咽腔內的構造與尾索動物相似,也具有內柱、咽上溝和圍咽溝等。文昌魚幼體的鰓裂直接開口於體表,後來形成圍鰓腔,以腹孔作為咽部鰓裂的總出水口。該攝食方式為被動濾食攝食。
咽內的食物微粒被內柱細胞的分泌物粘結成團,再由纖毛運動使它從後向前流動,經圍咽溝轉到咽上溝,往後推送進入腸內。腸為一直管,向前伸出一個盲囊,突入咽的右側,稱為肝盲囊(hepatic diverticulum),能分泌消化液,與脊椎動物的肝臟為同源器官。食物團中的小微粒可進入肝盲囊,被肝盲囊細胞所吞噬,營細胞內消化,大微粒在腸內分解成小微粒後,也轉到肝盲囊中進行細胞內消化,未消化的物質由肝盲囊重返腸中,在後腸部進行消化和吸收。腸的末端開口於身體左側的肛門。
咽腔是文昌魚完成呼吸作用的部位。咽壁兩側有60多對鰓裂,彼此以鰓條分開,鰓裂內壁布有纖毛上皮細胞和血管。水流進入口和咽時,藉纖毛上皮細胞的纖毛運動,通過鰓裂,並使之與血管內的血液進行氣體交換,最後,水再由圍鰓腔經腹孔排出體外。有人認為文昌魚纖薄的皮膚也具有直接從水中攝取氧氣的能力。
 脊索動物門
脊索動物門 5、血液循環文昌魚循環系統屬於閉管式,即血液完全在血管內流動,這種情形與脊椎動物基本相同。無心臟,但是具搏動能力的腹大動脈(ventral aorta),因而被稱為狹心動物。由腹大動脈往兩側分出許多成對的鰓動脈(branchial arteries)進入鰓隔,鰓動脈不再分為毛細血管,它在完成氣體交換的呼吸作用後,於鰓裂背部匯入2條背大動脈根。背大動脈根內含多氧血,往前流向身體前端,向後則由左、右背大動脈根合成背大動脈(dorsal aorta),再由此分出血管到身體各部。血液無色,也沒有血細胞和呼吸色素,動脈中的血液通過組織間隙進入靜脈。從身體前端返回的血液通過體壁靜脈(parietal vein)注入一對前主靜脈(anterior cardinal vein);尾的腹面有一條尾靜脈(caudal vein),收集一部分身體後部回來的血液,進入腸下靜脈(subintestinal vein),大部分血液則流進2條後主靜脈(posterior cardinal vein)。左、右前主靜脈和兩條後主靜脈的血液全部匯流至一對橫形的總主靜脈(common cardinal vein),或稱居維葉氏管(ducts Cuvieri)。左、右總主靜脈會合處為靜脈竇(sinus venosus),然後通入腹大動脈。從腸壁返回的血液由毛細血管網集合成腸下靜脈,尾靜脈的部分血液也注入其中;腸下靜脈前行至肝盲囊處血管又形成毛細管網,由於這條靜脈的兩端在肝盲囊區都形成毛細血管,因此稱作肝門靜脈(hepatic portal vein)。由肝門靜脈的毛細血管再一次合成肝靜脈(hepatic vein)並將血液匯入靜脈竇內。
脊椎動物
脊椎動物亞門(Vertebrates)
1、生活史某個時期具有脊索動物的三大特徵(其中咽鰓裂陸生脊椎動物出生後沒有)
2、神經系統發達,腦與脊髓分化,感覺器官發達,適應環境的能力更強
3、脊柱代替脊索成為支撐身體的中軸,更加堅固、靈活
4、具有心臟,加速血液循環
5、具有較完善的排泄器官(陸生動物比水生動物發達),更有效排除代謝廢物
6、具有成對附肢(圓口綱除外)擴大生活範圍,提高攝食、求偶、避敵的能力
脊椎動物亞門,脊索或多或少被脊柱所代替,腦和感覺器官集中於前端,形成明顯頭部,稱為有頭類(Craniata)。
下屬分類
尾索動物亞門
尾索動物亞門(Urochordata)
尾索動物是原始的特化的海棲脊索動物。幼體具上述3大特徵,但脊索在尾部。變態後脊索消失,背神經管退化成神經節,鰓裂仍存在。成體具被囊(tunic),大多營固著生活。
尾索動物和頭索動物兩個亞門是脊索動物中最低級的類群,總稱為原索動物(Protochorda-ta)。身體包在膠質(gelatinous)或近似植物纖維素成分的被囊中,至少在幼體時期尾部具有脊索及神經管,所以稱為尾索動物或被囊動物(tunicate)。全世界約有1370多種,常見種類有柄海鞘(Styela clava)、海樽(Ascida)、玻璃海鞘(Ciona)、菊花海鞘(Botryllus)等,分布遍及世界各地的海洋。體呈袋形或桶狀,包括單體或群體兩個類型,絕大多數無尾種類只在幼體時期自由生活,成體於淺海潮間帶營底棲固著生活,少數終生有尾種類在洋面上營漂浮式的自由游泳生活。體表有入水孔(incurrent siphon)和出水孔(excurrent siphon),咽壁有數量不等的鰓裂,咽外圍有寬大的圍鰓腔(peribranchial cavity),與出水管孔相通。一般為雌雄同體(hermaphroditism),異體受精。也有無性繁殖。由於卵和精子並不同時成熟,所以避免了自體受精。營有性生殖,也營無性的出芽生殖,樽海鞘的生活史中甚至還有複雜的世代交替現象。除個別種類外,受精卵都先 發育成善於游泳的蝌蚪狀幼體,再行變態發育(一般為逆行變態)。
循環系統是開管式,反向循環。
早在2千多年前,尾索動物就已經被記載和描述,曾先後隸屬於無脊椎動物中的蠕蟲類、擬軟體動物、苔蘚動物或軟體動物,直到1866年俄國學者柯瓦列夫斯基仔細地研究了海鞘的胚胎髮育及其變態後,才正式判定其正確的分類地位,置於脊索動物門。
柄海鞘
柄海鞘是尾索動物亞門中最主要的類群,約占全部種數的90%以上。柄海鞘是海鞘類中的優勢種,經常與盤管蟲(Hydroides)、藤壺(Balanus)及苔蘚蟲(Bugula)等附著在一起,固著在碼頭、船塢、船體,以及海水養殖的海帶筏和扇貝籠上,被作為沿海污損生物的重要指標種。柄海鞘的身體構造及變態過程在動物中具有一定的代表性,現以此為例簡述其主要特徵。
分類
本亞門是脊索動物中最低等的類群,遍布世界各個海洋,約1370多種,分屬於3綱,中國已知有14種左右。
尾海鞘綱
尾海鞘綱(Appendiculariae)本綱是尾索動物中的原始類型,共1目3科60餘種。體長數毫米至20mm,代表動物為住囊蟲(Oikopleura)和巨尾蟲(Megalocercus huxleyi)等。尾海鞘綱與本亞門中其他2綱的主要區別是:體外無被囊,只有兩個直接開口體外的鰓裂而缺乏國鰓腔,終生保持著帶有長尾的幼體狀態(neotonous),大多在沿岸淺海中營自由游泳生活。生長發育過程中無逆行變態,故又名幼形綱(Larva-cea)。住囊蟲包藏在由皮膚分泌的膠質住囊(gelatinous'house')內,住囊有入水孔和出水孔,住囊蟲在囊中藉助內有脊索和神經索的尾巴擺動進水,並使囊中的水由出水孔排出,推動動物體前進,同時通過蟲體口外特有的網篩(filter),從流水中濾取微 小的浮游生物作為食物。每隔數小時,住囊的出、入水孔就將被堵塞。此時住囊蟲即激烈揮動長尾,從特殊的應急出口(或稱小室孔道 escape hatch)破囊衝出至海中,並在很短的時間裡再形成新的住囊。
中國至今尚未發現本綱動物。
海鞘綱
海鞘綱(Ascidiacea)種類繁多,約有1250種,包括單體和群體2種類型,附著於水下物體或營水底固定生活。單體型種類的最大體長可達200餘毫米,群體的全長可超過0.5米以上。群體型種類的許多個體都以柄相連,並被包圍在一個共同的被囊內,但分別以各自的入水孔進水,有共同的排水口,如群體海鞘(Diplosoma)。
廣布於中國的海鞘綱動物有米氏小葉鞘(Leptoclinum mitsukurii)、星座美洲海鞘(Amarou-cium constellatum)、長紋海鞘(Ascidia longistriata)、玻璃海鞘(Ciona intestinalis)、3種菊海鞘(Botryllus ssp.)、瘤海鞘(Styela canops)、乳突皮海鞘(Molgula manhattensis)、龜甲海鞘(Chely-osoma)、西門登擬菊海鞘(Botrylloides simodensis)等。乳突皮海鞘和柄海鞘是連雲港以北黃、渤海沿岸污損生物中的優勢種,瘤海鞘大量出現在香港、西沙永興島和琛航島等南海海域內。
樽海鞘綱
樽海鞘綱(Thaliacea)本綱動物大多是營自由游泳生活的漂浮型海鞘,體呈桶形或樽形,咽壁有2個或更多的鰓裂。成體無尾,入水孔和出水孔分別位於身體的前後端。被囊薄而透明,囊外有環狀排列的肌肉帶,肌肉帶自前往後依次收縮時,流進入水孔的水流即可從體內通過出水孔排出,以此推動樽海鞘前進,並在此過程中完成攝食和呼吸作用。生活史較複雜,繁殖方式是有性與無性的世代交替。樽海鞘綱約有65種,代表動物有樽海鞘(Doliolum deuticula-tum),中國廈門沿海曾發現過小海樽(Dolioletta natilnalis)。磷海鞘(Pyrosoma atlanticum)為群體型種類,身體總長度可達200~600mm,因其口孔內緣有磷光器,漂浮游泳時能發出閃爍的磷光而得名,別名火體蟲。
頭索動物亞門
頭索動物亞門(Cephalochordata)
 文昌魚
文昌魚 沒真正的腦分化,故稱無頭類。
上述脊索動物的3大特徵終生存在。脊索縱貫全身,並向前延伸至背神經管前端。
頭索動物是一類終生具有發達脊索、背神經管和咽鰓裂等特徵的無頭魚形脊索動物。包括頭索綱(Cephalochorda,又名狹心綱Leptocardii)、鰓口科(Branchiostomidae),約30種,分隸於文昌魚和偏文昌魚(Asymmetron)2個屬。頭索動物分布很廣,遍及熱帶和溫帶的淺海海域,其中尤以北緯48°至南緯40°之間的沿海地區數量較多。
分類
頭索綱又稱狹心綱(Leptocardii),僅文昌魚目(Branchiostomiformes),包括文昌魚科(鰓口科)(Branchiostomidae)和偏文昌魚科(側殖文昌魚科)(Epiyonichthyidae)。文昌魚科生殖腺左右對稱排列於腹部兩側,有1屬9種,包括白氏文昌魚(Branchiostoma belcheri)、加州文昌魚(Branchiostoma californiense)、開普斯文昌魚(Branchiostoma capense)、卡氏文昌魚(Branchiostoma caribaeum)、佛州文昌魚(Branchiostoma floridae)、普通文昌魚(Branchiostoma lanceolatum)、瓦氏文昌魚(Branchiostoma valdiviae)、維吉尼文昌魚(Branchiostoma virginiae)。其中加州文昌魚產於北美聖地亞哥灣,長達100毫米,是體形最大者;我國最早發現的是廈門白氏文昌魚(Branchiostoma belcheri),廣泛分布於渤、黃、東、南海的淺水區。偏文昌魚科僅右側有一行生殖腺,有2屬5種,包括偏文昌魚屬(Asymmetron)和側殖文昌魚屬(Epigonichthys)。前者包括蘆卡偏文昌魚Asymmetron lucayanum和莫氏偏文昌魚Asymmetron maldivense,後者包括短刀側殖文昌魚Epigonichthys cultellus、蘆卡側殖文昌魚Epigonichthys lucayanus和馬爾地夫側殖文昌魚Epigonichthys maldivensis。我國有偏文昌魚(Asymmetron maldivense)和蘆卡側殖文昌魚(Epigonichthys lucayanus),分布於台灣南端的南灣水域。
脊椎動物亞門
該亞門包括:圓口綱、軟骨魚綱、硬骨魚綱、兩棲綱、爬行綱、鳥綱和哺乳綱。各綱的特徵雖然有顯著差別,但組成軀體的器官系統及其功能基本一致。
分類
 龜
龜 (1) 圓口綱(Cyclostomata):無頜,又稱無頜類(Agnatha);無成對附肢。脊索終生存在,並出現雛形脊椎骨。
(2) 甲冑魚綱:無頜,又稱無頜類(Agnatha),出現成對的鰭。脊索終身存在,並出現雛形脊椎骨。
(3) 盾皮魚綱:出現下頜骨,體表擁有骨板。
(4) 軟骨魚綱(Chondrichthyes)體被盾鱗,鰓裂直接開口於體外。
(5) 輻鰭魚綱(Osteichthyes)骨骼大多為硬骨,體被硬鱗、圓鱗或櫛鱗,鰓裂不直接開口於體表。
(6) 棘魚綱:背鰭、胸鰭、腹鰭和臀鰭的前端有硬棘。形體似鯊,歪形尾;胸、腹鰭發育完全,但鰭條不發育,在胸鰭和腹鰭之間有“額外”的偶鰭,或叫附加鰭;體被細小菱形鱗片,其結構似軟骨硬鱗魚;眼大,側生,前位並有圍眶骨;背鰭一個或兩個;有原始的頜,一個擴大的上頜骨與發育完善的下頜咬合,上頜無牙,下頜有牙;內骨骼已開始骨化。
(7) 肉鰭魚綱:擁有肉鰭,能夠初步上陸地。
(8) 兩棲綱(Amphibia):由水上陸的過渡種類,幼體魚形,以鰓呼吸,成體出現5指(趾)型四肢,皮膚裸露,以肺和皮膚呼吸。與其他更高等脊椎動物共稱為四足類(Tetrapoda)。
(5) 爬行綱(Reptilia):完全陸生。皮膚乾燥,被以角質鱗、角質骨片或骨板。肺呼吸。胚胎髮育中出現羊膜,與鳥類、哺乳類共稱為羊膜類(Amniotes)。其他各綱脊椎動物稱為無羊膜動物(Anomniotes)。
(6) 鳥綱(Aves):全身被羽,前肢變為翼,適應空中飛翔生活。血液循環為完全雙循環,恆溫,卵生。與哺乳類共稱為恆溫動物(Endotherm)。其他脊椎動物均為變溫動物(Ectotherm)。
(7) 哺乳綱(Mammalia):體外被毛,恆溫,胎生(單孔類除外),哺乳(具乳腺)。
生長發育
尾索動物亞門
柄海鞘成體的形態結構與典型的脊索動物有很大差異。然而,它的幼體外形酷似蝌蚪並具有脊索動物3個主要特徵。幼體長約0.5mm,尾內有發達的脊索,脊索背方有中空的背神經管,神經管的前端甚至還膨大成腦泡(cerebral vesicle),內含眼點和平衡器官等;消化道前段分化成咽,有少量成對的鰓裂;身體腹側有心臟。
體經過幾小時的自由生活後,就用身體前端的附著突起(adhesivepapillae)粘著在 其它物體上,開始其變態。在變態過程中,海鞘幼體的尾連同內部的脊索和尾肌逐漸萎縮,並被吸收而消失,神經管及感覺器官也退化而殘存為一個神經節。與此相反,咽部卻大為擴張,鰓裂數急劇增多,同時形成圍繞咽部的圍鰓腔;附著突起也為海鞘的柄所替代。附著突起背面因生長迅速,把口孔的位置推移到另一端(背部),於是造成內部器官的位置也隨之轉動了90°~180°的角度。最後,由體壁分泌被囊素構成保護身體的被囊,使它從自由生活的幼體變為營固著生活的柄海鞘。柄海鞘經過變態,失去了一些重要的構造,形體變得更為簡單,這種變態稱為逆行變態(retrogressive metamorphosis)。
頭索動物亞門
其典型代表種Branchostoma(文昌魚);體呈魚形,長40毫米左右,側扁,頭尾尖削,無頭與軀幹部之分,故名。體半透明,呈透明,呈微紅色,生活於淺海疏鬆的沙灘里,常鑽孔。以矽藻等為食。
頭索動物亞門(6張)
沿背側有一縱行皮膚褶皺,稱為背鰭;再後延至尾部高起,成為尾鰭,作矛形。自尾鰭腹面向前至體後約1/3處的皮膚褶襞,稱為腹褶(metapleure)。左右兩腹褶向後的匯合處是為腹孔,或圍鰓腔孔(abdominal pore),腹鰭與尾鰭之間的小孔,為肛門。頭部前端突出部位為吻。其後的腹面有一環形薄膜,成為口笠。其邊緣生有36-50條觸鬚。口笠的內腔稱為前庭。
神經管的形成:原腸胚背中線的外胚層細胞向內凹陷,形成神經板,其兩同時翹起形成一對縱褶,叫神經褶。神經褶又相向生長,在神經板背側癒合;神經板 兩側亦向上包圍生長;彼此接合。於是在背面出現一條中空的管道,即神經管。
脊索與中胚層的發生:在神經管形成的同時,原腸腔的背面、神經板下方的內胚層細胞向背方突出,逐漸脫離內胚層形成一條縱貫的脊索;在脊形成的同時,脊索突 起的兩側也發生若干成對的分節狀突起,最後脫離內胚層,自成一腔囊,叫體腔囊,也以是中胚層。將來發育成為體腔。內胚層除了分離成脊索和中胚層外,其他部分則形成腸管的內壁。
因具有縱貫體軀一直達到前端的脊索而得名;頭無明顯分化,與體軀不能截然分開,故又名無頭類,通稱文昌魚、矛形魚。頭索動物終生具有縱貫背部,起支撐作用的脊索;有背神經管和咽鰓裂。這些在高等脊索動物中只存在於胚胎或幼蟲期,成體消失或分化為更高級的器官。
文昌魚為雌雄異體。生殖腺附生於圍鰓腔兩側的內壁上,是26對左右厚壁的矩形小囊,性成熟時可根據文昌魚的精巢為白色或卵巢呈現淡黃色進行雌雄鑑別。成熟的精子和卵都是通過生殖腺壁的破口釋出,墜入圍鰓腔,再隨同水流由腹孔排出,在海水中完成受精作用。文昌魚在6~7月產卵,通常產卵和受精都在傍晚進行。卵小而含卵黃少,為均黃卵(isolecithal egg),卵徑0.1mm~0.2mm。
文昌魚的發育需經歷受精卵—桑椹胚—囊胚—原腸胚—神經胚各個時期,才孵化成幼體。受精卵進行幾乎均等的全分裂(holoblastic),經過多次細胞分裂後,使許多細胞結成一個形似實心圓球的桑椹胚(morula)。桑椹胚在繼續細胞分裂的同時,中心的細胞逐漸向胚體表面遷移,從而變成一個內部充滿膠狀液的空心囊胚(blastopore),囊胚中的腔為囊胚腔(blastocoel),囊胚上端的細胞略小,稱動物極(animal pole),細胞較大的下端是植物極(vegetative pole)。接著,囊胚的植物極大細胞向內陷入以至與上端動物極細胞的內壁互相緊貼,囊胚腔因受擠壓而消失,被新出現的原腸腔(archenteron)所代替。原腸腔以植物極細胞內陷處的胚孔(或稱原口 blas-topore)與外界相通,這裡相當於胚體的後端,相對的另一端為前端。此時胚胎已形成內、外兩層細胞,分別稱為內胚層(endoderm)和外胚層(ectoderm),胚體表面長有纖毛並能在胚膜中進行迴旋運動。胚胎髮育到這個階段稱為原腸胚(gastrula)。原腸胚的後期開始胚體延長、產生中樞神經、胚層進一步分化及各器官系統的形成。原腸胚自前端沿背中線至胚孔的外胚層下陷成神經板(neural plate),與此相鄰的兩側外胚層同神經板脫離,互相靠攏而完全癒合,是將來的表皮部。下陷到表皮內的神經板首先在板的兩側往上隆起成神經褶(neural fold),然後卷合圍成背面留有一條縱裂的神經管(neural tube),管內為神經管腔(neurocoel)。其前端以神經孔(neuropore)和外界相通,後端經胚孔與原腸相通成神經腸管(neurenteric canal)。成體時,神經孔關閉成嗅窩,而神經腸管也閉塞不通並在胚孔部形成肛門,此時的胚胎稱為神經胚(neurula)。在背神經管形成的同時,脊索和中胚層也在形成。原腸背面正中出現一條縱行的隆起實體,即脊索中胚層,它與原腸分離後發育成脊索。脊索兩側各有一列按節排列和彼此連線的體腔囊(coelomic sac),這就是新發生的中胚層,體腔囊中的每個空腔即體腔(coelom)。
文昌魚身體前部的中胚層是以體腔囊的方式所形成,與棘皮動物及半索動物相同,身體後部中胚層的發生方式又與脊椎動物一致,由此可見文昌魚在兩大類動物中處於過渡性質的居中地位,對動物學的一些基本理論研究方面,可以提供許多有價值的材料。隨著每個體腔囊的發育,又分化成背、腹兩部分。背部稱體節(somite),腹部稱側板(lateralplate),體節內的體腔以後自行消失,而側板內的體腔最初因體腔囊分節彼此獨立存在,後來由於體腔囊壁前後溝通,才在體內形成一個完整的體腔。這種體腔叫作次級體腔,是真正由中胚層所構成的體腔。體節的內側部分,又分化為生骨節(sclerotome),將來形成脊索鞘、背神經管外的結締組織和肌隔等;體節的中部形成肌節;體節的外側部分以後形成皮膚的真皮。側板的外層為體壁中胚層(somatic mesoderm),將來發育成緊貼著體腔壁的腹膜或體腔膜(peritoneum);內層稱髒壁中胚層(splanchnic mesoderm),以後形成腸管外圍的組織。髒壁中胚層在腸管前段的背側發生出分節排列的指狀突起,即未來的腎管。體壁與側板交界處的體腔壁上也發生突起,以後自此發育出文昌魚的生殖腺。受精卵經過20多個小時後,文昌魚的胚胎髮育基本結束,全身披有纖毛的幼體就能突破卵膜,到海水中活動,此時有白天游至海底夜間升上海面進行垂直洄游的生活規律。幼體期約3個月,然後沉落海底進行變態。幼體在生長發育和變態的過程中,身體日益長大,出現前庭,鰓裂的數目因發生次生鰓條而增加了一倍,並由原來直接開口體外而變為通入新形成的圍鰓腔中。一齡的文昌魚體長約40mm,性腺發育成熟,可參與當年的繁殖。
脊椎動物亞門
脊椎動物區別於其他脊索動物的特徵有:出現明顯的頭部,中樞神經系統成管狀,前端擴大為腦,其後方分化出脊髓。
大多數種類的脊索只見於發育早期(圓口綱、軟骨魚綱和硬骨魚綱例外),以後即為由單個的脊椎骨連線而成的脊柱所代替。原生水生動物用鰓呼吸,次生水生動物和陸棲動物只在胚胎期出現鰓裂,成體則用肺呼吸。除圓口綱外,都具備上、下頜。循環系統較完善,出現能收縮的心臟,促進血液循環,有利於提高生理機能。用構造複雜的腎臟代替簡單的腎管,提高排泄機能,由新陳代謝產生的大量廢物能更有效地排出體外。除圓口綱外,水生動物具偶鰭,陸生動物具成對的附肢。
生活習性
尾索動物亞門
柄海鞘的成體呈長橢圓形,幼體型似蝌蚪。基部以柄附生在海底或被海水淹沒的物體上,另一端有2個相距不遠的孔:頂端的一個是入水孔,孔內通消化管而中間有一片篩狀的緣膜,其作用是濾去粗大的物體,只容許水流和微小食物進入消化道;位置略低的一個是出水孔(圖14-3)。從胚胎髮生和幼體變態的過程來看,兩孔之間是柄海鞘的背部,對應的一側為腹部。一般情況下,水流從入水孔進入而由出水孔排出,當受驚擾或刺激時,則可引起體壁驟然收縮,體內積貯的水分別從2個孔中似乳汁般同時噴射而出,故在山東省沿海一帶俗稱海奶子;刺激緩解後,身體又逐漸恢復原狀。它們除了可以成簇密集生活外,還能附著在同種的其它個體上,同時本身又可以被別的個體所附著,呈現壘疊的聚生現象。
頭索動物亞門
頭索動物的脊索不但終生保留,且延伸至背神經管的前方,故稱頭索動物。又因本亞門動物都缺乏真正的頭和腦,所以又稱無頭類。中國廈門、青島等地所產的文昌魚(Branchiostomabelcheri),可作為頭索動物的代表。
白氏鰓口文昌魚(Branchiostoma belcheri)
文昌魚的體形略似小魚,無明顯的頭部,左右側扁,半透明,可隱約見到皮下的肌節
(myomere)和腹側塊狀的生殖腺;身體兩端尖出,故有雙尖魚(Amphioxus)之稱,又因其尾形很像矛頭而名海矛。一般體長約50mm,但產於美國的加州文昌魚(Branchiostomacaliforniense)可超過100mm,是該屬中已知個體最大的一種。前端的腹面為一漏斗狀的口笠(oral hood),口笠內為前庭(vestibule),內壁有輪器(wheel organ),由前庭引向位於一環形緣膜(velum)中央的口。口笠和緣膜的周圍分別環生觸鬚(cirri)及緣膜觸手(velar tentacle),具有保護和過濾作用,可阻擋粗砂等物隨水流進入口中。整個背面沿中線有一條低矮的背鰭(dorsalfin),往後與高而繞尾的尾鰭(caudal fin)相連。此外在肛門之前還有肛前鰭(preanal fin)。無偶鰭,只在身體前部的腹面兩側各有一條由皮膚下垂形成的縱褶,稱為腹褶(metapleura fold)。腹褶和肛前鰭的交界處有一腹孔(atripore),是咽鰓裂排水的總出口,故又名圍鰓腔孔。
脊椎動物亞門
脊椎動物數量多,結構複雜,進化地位高,形態結構彼此懸殊,生活方式千差萬別 ,不便一一介紹,詳見脊椎動物亞門。
演化意義
脊索的出現是動物演化史中的重大事件,使動物體的支持、保護和運動的功能獲得“質”的飛躍。這一先驅結構在脊椎動物達到更為完善的發展,從而成為在動物界中占統治地位的一個類群。
脊索(以及脊柱)構成支撐軀體的主梁,是體重的受力者,使內臟器官得到有力的支持和保護,運動肌肉獲得堅強的支點,在運動時不致由於肌肉的收縮而使軀體縮短或變形,因而有可能向“大型化”發展。脊索的中軸支撐作用也使動物體更有效地完成定向運動,對於主動捕食及逃避敵害都更為準確、迅捷。脊椎動物頭骨的形成、頜的出現以及椎管對中樞神經的保護,都是在此基礎上進一步完善化的發展。
頭索動物
頭索動物身體結構比較簡單,但脊索動物的典型結構已經具有,三大結構終生存在。根據形態學推測,脊索動物由一種左右不對稱,無圍腮腔、腮裂少、營自由生活的原始無頭類演化而來,且在進化當中分為兩支,一支適應自由生活,演變為原始有頭類,進而走向脊椎動物;另一支向底棲生活發展,演變為尾索動物和頭索動物。
尾索動物
尾索動物是最低等的脊索動物,與高等脊索動物存在著演化上的親緣關係,兩者可能都是從類似海鞘幼蟲型營自由生活的共同祖先--原始無頭類動物演化而來。這類原始無頭類動物不但將幼體時期的尾和自由游泳的生活方式保留到成體,甚至還消失了生活史中營固著生活的階段,並通過幼態滯留及幼體性成熟途徑發展為頭索動物和脊椎動物。尾索動物是在進化過程中適應特殊生活方式的一個退化分支,除保留濾食的咽及營呼吸作用的咽鰓裂外,大多數種類已在變態中失去所有的進步特徵,並向固著生活的方向發展。
脊椎動物
脊椎動物亞門(Vertebrata)是動物界中數量最多,結構最複雜,進化地位最高的類群。形態結構彼此懸殊,生活方式千差萬別。
脊椎動物的脊索只在胚胎髮育階段出現,隨後或多或少地被脊柱所代替腦和各種感覺器官在前段集中,形成明顯的頭部,故稱有頭類。
生物區別
與無脊椎動物
①脊索:脊索動物具有縱貫背部的脊索,後被脊柱所代替;無脊椎動物無脊索或脊柱。
②中樞神經:脊索動物中空的神經中樞位於背部;無脊椎動物原生與海綿動物無神經系統,腔腸動物為網狀神經系統,扁蟲與線蟲為梯狀神經系統,環節與節肢動物中樞神經呈索狀位於身體腹面。
③鰓裂:脊索動物生活史的全部或部分時期具有鰓裂;無脊椎動物不具鰓裂。
④心臟位置:脊索動物心臟位於消化道腹面;無脊椎動物心臟位於消化道背面或無心臟。
⑤生物化學方面:脊索動物參與肌肉收縮能量代謝的非蛋白質含氮浸出物是磷酸肌酸;無脊椎動物的是磷酸精氨酸。
脊索動物除上述特徵外,其一些結構也見於一些無脊椎動物中,如後口、三胚層、兩側對稱、真體腔、分節現象、閉管式血液循環等。兩者的結構比較見無脊椎動物。
