
人種簡介
具有共同遺傳體質特徵的人類群體。人種也稱種族,在生物學上,人類各種族都同屬於一個物種,即智人。不同的種族相當於在一個物種下的若干變種,他們都起源於一共同祖先。不同的人種雖然在膚色、眼色、發色、髮型、頭型、身高等特徵上有所區別,但這些特徵差異是由於人類在一定地域內長期適應當地自然環境,又經長期隔離所形成的。分類
根據體質特徵的差異,過去全世界人種大致上分為3大人種,即蒙古人種(黃色人種)、高加索人種(白色人種)、尼格羅人種(黑色人種)。有人還主張再分出澳大利亞人種(棕色人種)來。在這些主要人種之間還有若干過渡人種,如介於尼格羅人種和高加索人種之間的有衣索比亞人種和南印度人種,介於高加索人種和蒙古人種之間的有南西伯利亞人種和烏拉爾人種,介於蒙古人種和尼格羅人種之間的有玻里尼西亞人種和千島人種。隨著人類歷史的發展,人類的生產活動能力的提高,控制自然能力的增強,社會因素逐漸加強而自然選擇因素逐漸削弱,人類交往日益頻繁,各人類集團間不斷發生混雜,混雜現象越來越廣泛,幾乎擴展到了世界上每個角落,因此,世界上幾乎沒有一個絕對的純種。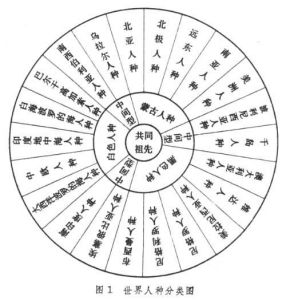 人種
人種現代各人種的分布
除去各種水域和一些高山外,人類是在地球上分布最廣的一種生物。現代人的分布經歷了一個相當長時期的發展,至少可以追溯到更新世晚期的後半部,即舊石器時代晚期的晚期智人階段。晚期智人具有高而圓隆的頭顱和相對較小的面部,低矮的眉弓和突出的下頦等特徵,它是現代智人的一種早期類型。在地球上廣泛分布的最早的晚期智人各群體已經積累了文化上和體質遺傳上的差異,並在此後3.5萬年以上的時間裡進一步得到擴展和改變,形成了現生各個人種的特徵和分布狀況。
與動物物種產生和分化的過程一樣,形成現代各人種的自然過程仍然受基因突變、變異的隨機擴散、遷移和自然選擇四個基本因素控制。在這些因素作用下,現生各人種在某些形態特徵和某些生物化學特徵上顯示了程度不同的差異,上述生物化學特徵包括ABO、Rh、MN等多種血型系統和其他血液因子系統,某些遺傳性疾病的存在或出現頻率,以及對某些疾病的遺傳性抗禦等等。
各人種的體質形態一般來說與他們的生活環境是相適應的。例如,居住在炎熱的非洲沙漠的尼格羅人的肢體細長;而在嚴寒的北極生活的因紐特人(以前一般稱為“愛斯基摩人”)則肢體粗短。這與身體熱量的保持和散發有關。赤道附近尼格羅人的皮膚含有較多的色素,可以避免過多的紫外線照射,他們捲曲的頭髮也可起隔熱的作用。蒙古人的傾斜的鳳眼和內眥褶,還有寬的和富含脂肪的面頰、較平的鼻部和額部,可能與亞洲中部寒冷的多風沙氣候有關。
在人種形成的過程中,自然因素作用的大小、性質和誘發條件,隨著人類物質生產和社會文化的發展而變得越來越不同於其他動物物種形成的過程。人種的形成和分布比在其他動物同一物種內不同亞種的情況複雜得多。因此,不能簡單地用自然條件來解釋某些人種的某些特徵。雖然非洲尼格羅人皮膚黑色素的驟增與太陽照射有關,但是實際上現在的非洲尼格羅人大部居住在蔽蔭的森林裡。世界上個子最高的人是居住在蘇丹尼羅河上游的尼羅特人,成年男子平均身高達180厘米。有趣的是,世界上最矮的俾格米人也居住在這個區域。斯堪的納維亞半島低矮的拉普人也與高大的北歐人種毗鄰居住,非洲尼格羅人面部突頜的特徵適宜於炎熱的氣候,但北極人種中的阿留申類型的突頜比尼格羅人還要明顯。這樣一些表面上與自然條件相矛盾的人種特徵的形成過程更複雜一些。
各人種的起源
蒙古人種、高加索人種、尼格羅人種三大人種主幹的體質特徵是在約 3.5萬年前開始出現的。在亞、歐、非三大洲發現的晚期智人的化石證明他們在本地區延續發展。美洲、澳洲和大洋洲較晚才有人類居住。由於澳洲土著的體質特徵較為特殊,他們常被列為與以上三種人不同的另一種人。蒙古人種起源於中亞和東亞。中國北京周口店山頂洞和廣西柳江縣通天岩發現的晚期智人頭骨化石表現了以黃種人特徵為主導的特徵。在馬來西亞沙撈越的尼亞洞和在印度尼西亞爪哇瓦賈克發現的人類化石很像現代澳大利亞土著。早期蒙古人種逐漸向北方擴展,成為西伯利亞的通古斯人和楚克奇人以及因紐特人(愛斯基摩人)。跨過西伯利亞和美洲阿拉斯加之間曾經存在的“陸橋”,蒙古人種來到美洲,定居下來,成為美洲大陸最早的居民──印第安人。
蒙古人種從印度尼西亞,或者還有其他的地方,航海到澳大利亞,比到美洲去的歷史或許稍早一些。在澳大利亞的芒戈湖、科阿沼澤發現的人類化石代表了兩種不同的類型,因此有人認為曾經有過兩次從亞洲向澳洲的遷移。大洋洲的美拉尼西亞、玻里尼西亞和密克羅尼西亞可能只是在幾千年前才有人居住。
非洲尼格羅人與在本地區發現的晚期智人在體質特徵上的繼承關係很明顯,大部分人認為他們起源於非洲本土。但是,地區內變種分化的較晚。南非和東非的晚期智人化石都在不同程度上包含尼格羅人 (Negroes)、科伊桑人(Khoisans,即以前所稱的“布須曼人和霍屯督人”)和當地現代人特徵的混合。只是在幾千年之前,科伊桑人才在非洲南部定居下來,在北部,與狩獵經濟相適應,形成尼格羅類形。
與歐洲晚期智人有關係的早期智人,即尼安德特人(Neandertals) 階段的化石比較複雜。最早的白種人的起源地目前只能圈定在南歐、北非和西亞這樣一塊較大的區域之中。歐洲本土發現的晚期智人化石以克羅馬農人(Cro-Magnon)為代表,他們在3萬年前已經顯示了高加索人種的性質。由於歐洲與附近亞、非洲部分的人群混雜很頻繁,高加索人種再分化為現代的次級人種則是大約 1萬年以內的事。這樣的次級人種包括波羅的海周圍淺膚色的北歐人種、淺黑色皮膚的地中海人種和北非衣索比亞人種以及深褐色皮膚的南印度人種。1000多年以前宗教對婚姻的限制也是後期人種特徵相對穩定的重要因素。
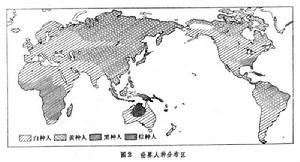
義大利航海家C.哥倫布1492年到達美洲大陸以後,隨著歐洲資本主義國家尋求新的殖民地,世界人種的分布發生了急劇的變化:數以百萬計的歐洲人湧向美洲、南非、澳大利亞和紐西蘭,使高加索人種的數目迅速增長。例如,近4個世紀以來世界人口增長約6倍,而英國人的後裔至少增長50倍。與此同時,幾百萬非洲黑人作為奴隸被綁架到美洲。由於混雜又出現了新的人種,美洲的有色人種已達6000萬人之多。有的人種(塔斯馬尼亞人)已滅絕,有些人種如澳洲土著及美洲印第安人人口大大減少。這樣的一系列過程形成了現代人種分布的基本格局(圖2)。
 人種
人種人種劃分的補充說明
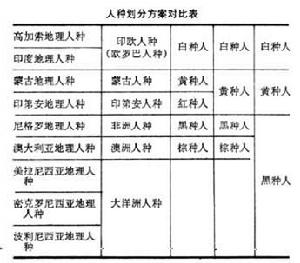
除外表形態特徵外,在生物化學及人類遺傳學研究的基礎上,發現過去以純形態學為標準的三分或四分的人種劃分方案已經不能全面反映真實情況了。至於完全以膚色為標準的“白種人”、“黃種人”、“黑種人”、“棕種人”,甚至還有“紅種人”的名稱也並不十分恰當,應該摒棄。基於科學的發展,在外表形態特徵基礎上,加上血型、遺傳病等的差異,再考慮地理阻障等因素,把世界人種劃分為9個地理人種(geographical race)(見表)。
 人種
人種高加索地理人種的Rh陰性血型出現率高,並有A2型血型的人,與印度地理人種不同。其膚色淺、體毛濃密。鼻大而窄和男子禿頂者多也是其顯著特徵。
印第安地理人種或稱美洲印第安地理人種,過去曾依外表特徵而歸於蒙古人種,如直黑髮、鏟形門齒出現率高、體毛稀疏、男子禿頂者少等。但其膚色較蒙古人種更濃重,且具偏紅色調。其血型特徵也與蒙古人種不同,如無B型及Rh陰性血型的人,N型血型的人也很少,而Diego陽性血型出現率高。
蒙古地理人種在血型特徵上是 B型血型出現率高,Duffy血型及Diego陽性血型出現率不穩定或偏高。
尼格羅地理人種從外形特徵上有時可以劃分為具典型特徵的剛果人種和膚色微黃,身材矮細、具螺鏇狀叢生髮的開普人種(包括科伊桑人)。二者在血型上無明顯差異。一般R0型血型出現率高,U-陰性血型略少,Fy型和V型血型很罕見。血紅蛋白異常。有6-磷酸葡萄糖脫氫酶缺乏症。
澳大利亞地理人種膚色濃黑,發波狀至捲曲狀,身材細長、體毛中等至濃密、男子禿頂率高、並伴有小腿無毛現象。牙齒大、眉嵴高、下頜突出。B型血型出現率很低,N型血型出現率極高,無S型血型。無異常血紅蛋白,無6-磷酸葡萄糖脫氫酶缺乏症,與尼格羅人種相反。
大洋洲諸島上的人種曾與澳洲人種共稱澳洲大洋洲人種。美拉尼西亞地理人種外貌上近似澳大利亞人種,但鏟形門齒出現率高,B型和S型血型出現率高。有6-磷酸葡萄糖脫氫酶缺乏症和地中海貧血基因。可能澳大利亞人種與之關係更密切些。
密克羅尼西亞地理人種曾被認為可能是東南亞蒙古人種、美拉尼西亞人種和玻里尼西亞人種的混血後裔。但其B型血型出現率稍高、N型較M型血型多、Duffy-陽性和Diego-陰性血型出現率很高。表明他們是獨立的地理人種。
玻里尼西亞地理人種膚色較淺、體毛少。B型血型出現率低、N型血型出現率較高,無Rh-陰性和A2型血型,B型、M型和Duffy-陽性血型出現率也不高。雖然面部特徵近似蒙古人種;但膚色淺,波狀發和濃密絡腮鬍又與高加索人種近似。其血型與蒙古人種及高加索人種均有較大差異。
在地理人種範圍內,基於生物學的或其他如地理阻障、社會、民俗等因素,甚至歷史和政治因素等而形成一些稍小的居群集團,可以稱之為地域人種(locol race)。他們往往只在本群內通婚,極少與其他集團交往。一個地域人種內的各成員間,生物學特徵極相似。往往許多在遺傳特徵上相似的地域人種可以在一個大陸或某一較大區域構成地理人種。處於邊緣地區、地理阻隔嚴重、人口很少的地域人種常常構成在遺傳性質上穩定的特殊的居群。
在人口稠密或地理阻限不嚴重的經濟發達地區,則諸多地域人種間生物學上的差異較少,而政治、歷史等方面的差異起作用較大。例如高加索人種中就有阿爾卑斯、地中海、東北歐、西北歐等地域人種。日本的阿伊努人、非洲的俾格米人、夏威夷群島的夏威夷人都屬於特定的地域人種。
在人數較多,人口密度較大而又少流動的地方常有小的居群集團;他們常表現為具單獨的遺傳因素,目前名之為少數人種 (microrace)。斯堪的納維亞的拉普人可作為少數人種的例子。
人類的種族(Races of Mankind)這一概念使用已久,目前常把人種寫成(human population),也就是賦予人種以居群意義。
人種的起源、形成、分布和混雜等問題至今仍有許多地方無法弄清。有待今後深入研究。
起源
除去各種水域和一些高山外,人類是在地球上分布最廣的一種生物。
現代人的分布經歷了一個相當長時期的發展,至少可以追溯到更新世晚期的後半部,即舊石器時代晚期的晚期智人階段。晚期智人具有高而圓隆的頭顱和相對較小的面部,低矮的眉弓和突出的下頦等特徵,它是現代智人的一種早期類型。在地球上廣泛分布的最早的晚期智人各群體已經積累了文化上和體質遺傳上的差異,並在此後3.5萬年以上的時間裡進一步得到擴展和改變,形成了現生各個人種的特徵和分布狀況。
與動物物種產生和分化的過程一樣,形成現代各人種的自然過程仍然受基因突變、變異的隨機擴散、遷移和自然選擇四個基本因素控制。在這些因素作用下,現生各人種在某些形態特徵和某些生物化學特徵上顯示了程度不同的差異,上述生物化學特徵包括ABO、Rh、MN等多種血型系統和其他血液因子系統,某些遺傳性疾病的存在或出現頻率,以及對某些疾病的遺傳性抗禦等等。
各人種的體質形態一般來說與他們的生活環境是相適應的。例如,居住在炎熱的非洲沙漠的尼格羅人的肢體細長;而在嚴寒的北極生活的因紐特人(以前一般稱為“愛斯基摩人”)則肢體粗短。這與身體熱量的保持和散發有關。赤道附近尼格羅人的皮膚含有較多的色素,可以避免過多的紫外線照射,他們捲曲的頭髮也可起隔熱的作用。蒙古人的傾斜的鳳眼和內眥褶,還有寬的和富含脂肪的面頰、較平的鼻部和額部,可能與亞洲中部寒冷的多風沙氣候有關。
分布
蒙古人種、高加索人種、尼格羅人種三大人種主幹的體質特徵是在約 3.5萬年前開始出現的。在亞、歐、非三大洲發現的晚期智人的化石證明他們在本地區延續發展。美洲、澳洲和大洋洲較晚才有人類居住。由於澳洲土著的體質特徵較為特殊,他們常被列為與以上三種人不同的另一種人。
蒙古人種起源於中亞和東亞。中國北京周口店山頂洞和廣西柳江縣通天岩發現的晚期智人頭骨化石表現了以黃種人特徵為主導的特徵。在馬來西亞沙撈越的尼亞洞和在印度尼西亞爪哇瓦賈克發現的人類化石很像現代澳大利亞土著。早期蒙古人種逐漸向北方擴展,成為西伯利亞的通古斯人和楚克奇人以及因紐特人(愛斯基摩人)。跨過西伯利亞和美洲阿拉斯加之間曾經存在的“陸橋”,蒙古人種來到美洲,定居下來,成為美洲大陸最早的居民──印第安人。
蒙古人種從印度尼西亞,或者還有其他的地方,航海到澳大利亞,比到美洲去的歷史或許稍早一些。在澳大利亞的芒戈湖、科阿沼澤發現的人類化石代表了兩種不同的類型,因此有人認為曾經有過兩次從亞洲向澳洲的遷移。大洋洲的美拉尼西亞、玻里尼西亞和密克羅尼西亞可能只是在幾千年前才有人居住。
非洲尼格羅人與在本地區發現的晚期智人在體質特徵上的繼承關係很明顯,大部分人認為他們起源於非洲本土。但是,地區內變種分化的較晚。南非和東非的晚期智人化石都在不同程度上包含尼格羅人 (Negroes)、科伊桑人(Khoisans,即以前所稱的“布須曼人和霍屯督人”)和當地現代人特徵的混合。只是在幾千年之前,科伊桑人才在非洲南部定居下來,在北部,與狩獵經濟相適應,形成尼格羅類形。 與歐洲晚期智人有關係的早期智人,即尼安德特人(Neandertals) 階段的化石比較複雜。最早的白種人的起源地目前只能圈定在南歐、北非和西亞這樣一塊較大的區域之中。歐洲本土發現的晚期智人化石以克羅馬農人(Cro-Magnon)為代表,他們在3萬年前已經顯示了高加索人種的性質。由於歐洲與附近亞、非洲部分的人群混雜很頻繁,高加索人種再分化為現代的次級人種則是大約 1萬年以內的事。這樣的次級人種包括波羅的海周圍淺膚色的北歐人種、淺黑色皮膚的地中海人種和北非衣索比亞人種以及深褐色皮膚的南印度人種。1000多年以前宗教對婚姻的限制也是後期人種特徵相對穩定的重要因素。
義大利航海家C.哥倫布1492年到達美洲大陸以後,隨著歐洲資本主義國家尋求新的殖民地,世界人種的分布發生了急劇的變化:數以百萬計的歐洲人湧向美洲、南非、澳大利亞和紐西蘭,使高加索人種的數目迅速增長。例如,近4個世紀以來世界人口增長約6倍,而英國人的後裔至少增長50倍。與此同時,幾百萬非洲黑人作為奴隸被綁架到美洲。由於混雜又出現了新的人種,美洲的有色人種已達6000萬人之多。
差異
英文:appearancedifferenceofhumanrace
釋文:不同人種在體質形態上有許多差異。
膚色:膚色有白色、黃色、褐色、棕紅色和黑色等,詳細劃分可分為32種不同色調。
眼色:眼色由虹膜中黑色素分布區域和數量多少決定,有黑褐色、褐色、淺綠色、灰色、天藍色等,並可詳細劃分為12~16級。
發色:發色包括灰金黃色、淺金黃色、金黃色、深金黃色14種黃色調;棕色有6種色調;黑棕色及火紅色各有6種色調,發色共有32種色調,有專門的帶實物的發色表可供對比。
●頭髮的形狀可分為直發、波狀發、捲髮三大類,其下又分為硬直、平直、微波、寬波、窄波、微卷、松卷、緊卷、松螺鏇卷和緊螺鏇卷等。
●髭鬚可根據多少分為五級;
○極少(髭鬚不相接、耳前極少); ○少(髭鬚在口角相連、耳前至下頜角亦有但不與下頜的須相連);○中等(髭鬚相連、兩側耳前連線成帶但面頰部極少);○多(髭鬚相連但頰部不濃密);○極多(十分濃密包括顴骨以下的面頰部)。
鼻部:鼻部包括鼻樑、鼻尖的形態和鼻翼部的寬窄和突度等。
唇:唇可根據上唇皮膚部的突度、紅唇的厚薄和外翻程度等區分為多種類型。
頭形:頭形(顱形)是從頂面觀察時顱骨的形態,大體上前後為長、左右為寬,可有長頭型(顱寬顱長比小於70)、短頭型(顱寬顱長比大於80)等7個級別。
面形:面形可形象化的分為方形、倒梯形、圓形、橢圓形等10種類型,亦可根據面高(鼻根至頦下)和面寬(兩顴骨最高點間)之比分為寬面型與窄面型等五級。
此外如蒙古人種常具內眥褶(蒙古褶)、鏟狀門齒和新生兒骶部的色素沉積(青色的蒙古斑);常見於尼格羅人種及太平洋人種的臀部脂肪聚積及外生殖器、乳房的特徵。
